

Modulation of GABAA receptors in cerebellar granule neurons by ethanol: a review of genetic and electrophysiological studies
- PMID: 17521847
- PMCID: PMC1986723
- DOI: 10.1016/j.alcohol.2007.04.004
Modulation of GABAA receptors in cerebellar granule neurons by ethanol: a review of genetic and electrophysiological studies
Abstract
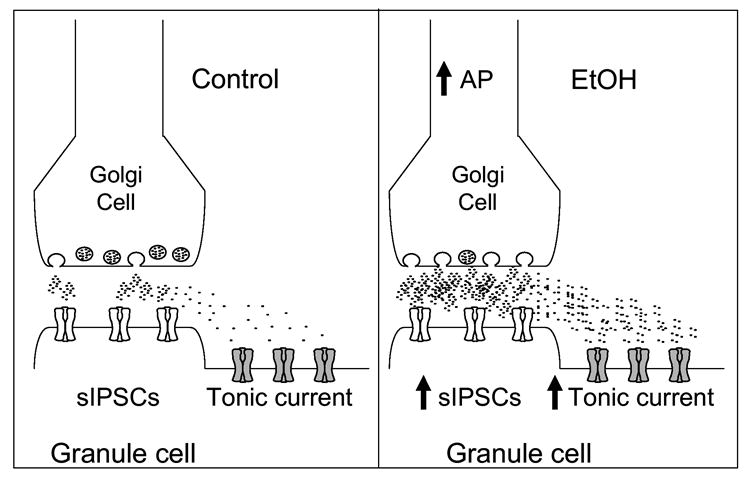
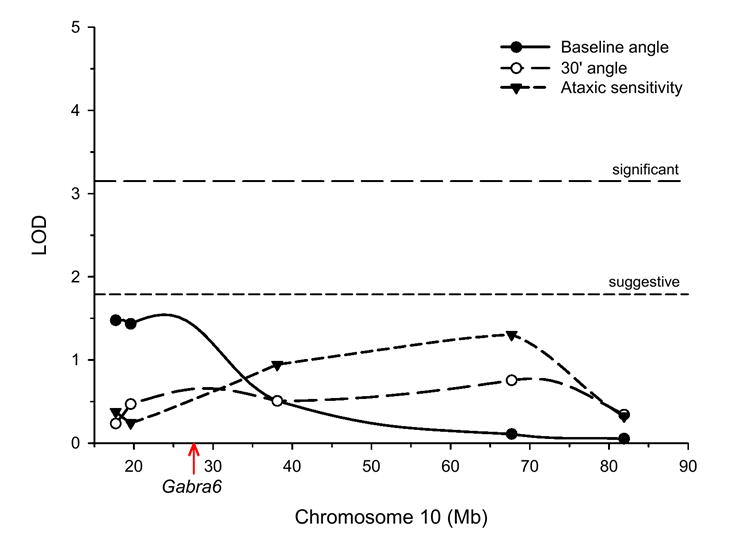
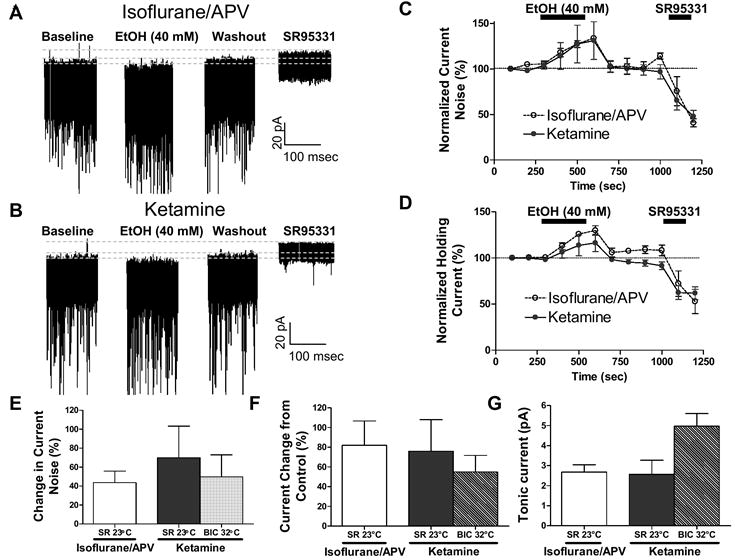
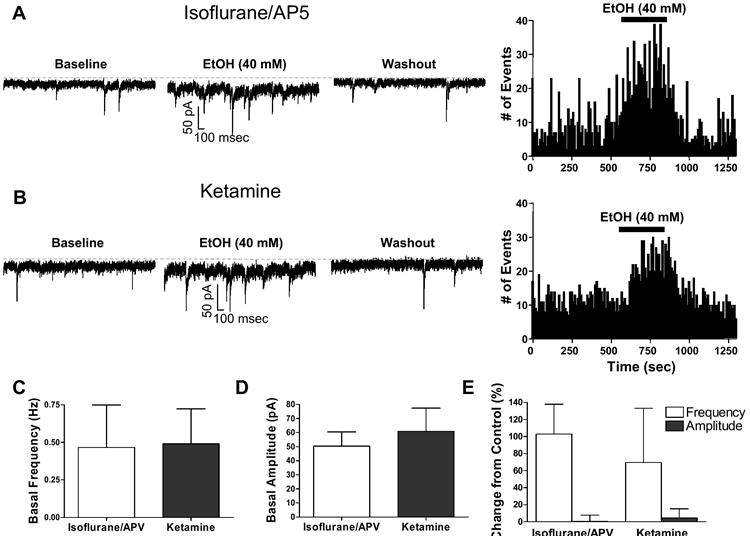
Cerebellar granule neurons (CGNs) receive inhibitory input from Golgi cells in the form of phasic and tonic currents that are mediated by postsynaptic and extrasynaptic gamma-aminobutyric acid type A (GABAA) receptors, respectively. Extrasynaptic receptors are thought to contain alpha6betaxdelta subunits. Here, we review studies on ethanol (EtOH) modulation of these receptors, which have yielded contradictory results. Although studies with recombinant receptors expressed in Xenopus oocytes indicate that alpha6beta3delta receptors are potently enhanced by acute exposure to low (>or=3 mM) EtOH concentrations, this effect was not observed when these receptors were expressed in Chinese hamster ovary cells. Slice recordings of CGNs have consistently shown that EtOH increases the frequency of phasic spontaneous inhibitory postsynaptic currents (sIPSCs), as well as the tonic current amplitude and noise. However, there is a lack of consensus as to whether EtOH directly acts on extrasynaptic receptors or modulates them indirectly; that is, via an increase in spillover of synaptically released GABA. It was recently demonstrated that an R to Q mutation of amino acid 100 of the alpha6 subunit increases the effect of EtOH on both sIPSCs and tonic current. These electrophysiological findings have not been reproducible in our hands. Moreover, it was shown the alpha6-R100Q mutation enhances sensitivity to the motor-impairing effects of EtOH in outbred Sprague-Dawley rats, but this was not observed in a line of rats selectively bred for high sensitivity to EtOH-induced motor alterations (Alcohol Non-Tolerant rats). We conclude that currently there is insufficient evidence conclusively supporting a direct potentiation of extrasynaptic GABAA receptors following acute EtOH exposure in CGNs.
Figures
Similar articles
-
Mini-Review: Effects of Ethanol on GABAA Receptor-Mediated Neurotransmission in the Cerebellar Cortex--Recent Advances.Cerebellum. 2015 Aug;14(4):438-46. doi: 10.1007/s12311-014-0639-3. Cerebellum. 2015. PMID: 25575727 Review.
-
Sensitivity of GABAergic Tonic Currents to Acute Ethanol in Cerebellar Granule Neurons is Not Age- or δ Subunit-Dependent in Developing Rats.Alcohol Clin Exp Res. 2016 Jan;40(1):83-92. doi: 10.1111/acer.12940. Alcohol Clin Exp Res. 2016. PMID: 26727526 Free PMC article.
-
Ethanol sensitivity of GABAergic currents in cerebellar granule neurons is not increased by a single amino acid change (R100Q) in the alpha6 GABAA receptor subunit.J Pharmacol Exp Ther. 2007 Nov;323(2):684-91. doi: 10.1124/jpet.107.127894. Epub 2007 Aug 17. J Pharmacol Exp Ther. 2007. PMID: 17704355
-
Recreational concentrations of alcohol enhance synaptic inhibition of cerebellar unipolar brush cells via pre- and postsynaptic mechanisms.J Neurophysiol. 2017 Jul 1;118(1):267-279. doi: 10.1152/jn.00963.2016. Epub 2017 Apr 5. J Neurophysiol. 2017. PMID: 28381493 Free PMC article.
-
GABAA receptor subtypes: the "one glass of wine" receptors.Alcohol. 2007 May;41(3):201-9. doi: 10.1016/j.alcohol.2007.04.006. Alcohol. 2007. PMID: 17591543 Free PMC article. Review.
Cited by
-
The Cerebellar GABAAR System as a Potential Target for Treating Alcohol Use Disorder.Handb Exp Pharmacol. 2018;248:113-156. doi: 10.1007/164_2018_109. Handb Exp Pharmacol. 2018. PMID: 29736774 Free PMC article.
-
Novel subunit-specific tonic GABA currents and differential effects of ethanol in the central amygdala of CRF receptor-1 reporter mice.J Neurosci. 2013 Feb 20;33(8):3284-98. doi: 10.1523/JNEUROSCI.2490-12.2013. J Neurosci. 2013. PMID: 23426657 Free PMC article.
-
Ethanol modulates facial stimulation-evoked outward currents in cerebellar Purkinje cells in vivo in mice.Sci Rep. 2016 Aug 4;6:30857. doi: 10.1038/srep30857. Sci Rep. 2016. PMID: 27489024 Free PMC article.
-
Mini-Review: Effects of Ethanol on GABAA Receptor-Mediated Neurotransmission in the Cerebellar Cortex--Recent Advances.Cerebellum. 2015 Aug;14(4):438-46. doi: 10.1007/s12311-014-0639-3. Cerebellum. 2015. PMID: 25575727 Review.
-
Potentiation of Gamma Aminobutyric Acid Receptors (GABAAR) by Ethanol: How Are Inhibitory Receptors Affected?Front Cell Neurosci. 2016 May 6;10:114. doi: 10.3389/fncel.2016.00114. eCollection 2016. Front Cell Neurosci. 2016. PMID: 27199667 Free PMC article. Review.
References
-
- Allan AM, Harris RA. Gamma-aminobutyric acid and alcohol actions: neurochemical studies of long sleep and short sleep mice. Life Sci. 1986;39:2005–2015. - PubMed
-
- Attwell D, Barbour B, Szatkowski M. Nonvesicular release of neurotransmitter. Neuron. 1993;11:401–407. - PubMed
-
- Basile A, Hoffer B, Dunwiddie T. Differential sensitivity of cerebellar purkinje neurons to ethanol in selectively outbred lines of mice: maintenance in vitro independent of synaptic transmission. Brain Res. 1983;264:69–78. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
