

Akt1 governs breast cancer progression in vivo
- PMID: 17460049
- PMCID: PMC1863437
- DOI: 10.1073/pnas.0605874104
Akt1 governs breast cancer progression in vivo
Abstract
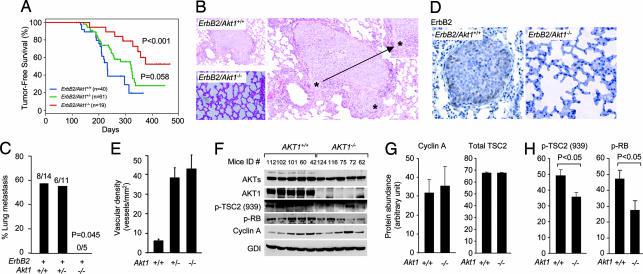
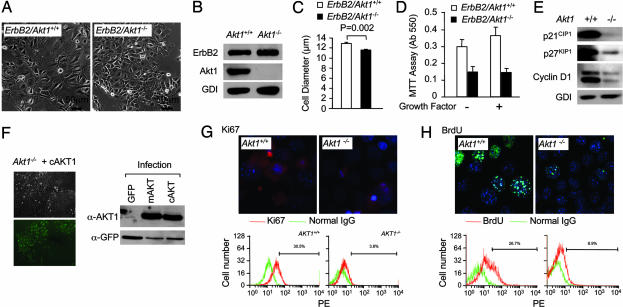
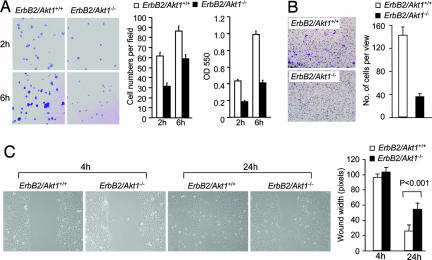
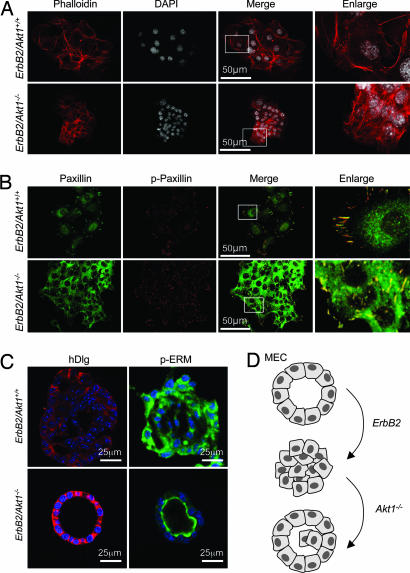
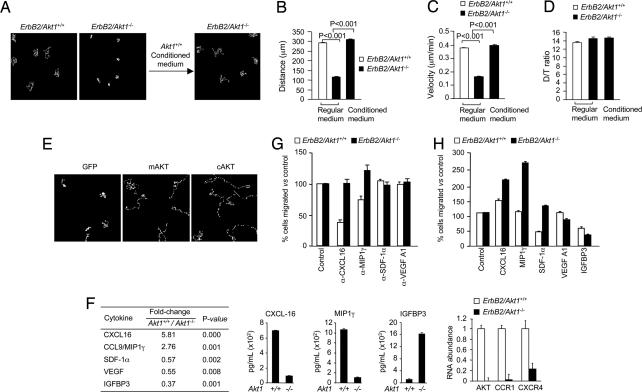
The serine threonine kinase Akt1 has been implicated in the control of cellular metabolism, survival and growth. Here, disruption of the ubiquitously expressed member of the Akt family of genes, Akt1, in the mouse demonstrates a requirement for Akt1 in ErbB2-induced mammary tumorigenesis. Akt1 deficiency delayed tumor growth and reduced lung metastases, correlating with a reduction in phosphorylation of the Akt1 target, tuberous sclerosis 2 (TSC2) at Ser-939. Akt1-deficient mammary epithelial tumor cells (MEC) were reduced in size and proliferative capacity, with reduced cyclin D1 and p27(KIP1) abundance. Akt1 deficiency abrogated the oncogene-induced changes in polarization of MEC in three-dimensional culture and reverted oncogene-induced relocalization of the phosphorylated ezrin-radixin-moesin proteins. Akt1 increased MEC migration across an endothelial cell barrier, enhancing the persistence of migratory directionality. An unbiased proteomic analysis demonstrated Akt1 mediated MEC migration through paracrine signaling via induction of expression and secretion of CXCL16 and MIP1gamma. Akt1 governs MEC polarity, migratory directionality and breast cancer onset induced by ErbB2 in vivo.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Akt1 ablation inhibits, whereas Akt2 ablation accelerates, the development of mammary adenocarcinomas in mouse mammary tumor virus (MMTV)-ErbB2/neu and MMTV-polyoma middle T transgenic mice.Cancer Res. 2007 Jan 1;67(1):167-77. doi: 10.1158/0008-5472.CAN-06-3782. Cancer Res. 2007. PMID: 17210696
-
AKT1 and AKT2 isoforms play distinct roles during breast cancer progression through the regulation of specific downstream proteins.Sci Rep. 2017 Mar 13;7:44244. doi: 10.1038/srep44244. Sci Rep. 2017. PMID: 28287129 Free PMC article.
-
ErbB2/neu kinase modulates cellular p27(Kip1) and cyclin D1 through multiple signaling pathways.Cancer Res. 2001 Sep 1;61(17):6583-91. Cancer Res. 2001. PMID: 11522658
-
miR-17/20 sensitization of breast cancer cells to chemotherapy-induced apoptosis requires Akt1.Oncotarget. 2014 Feb 28;5(4):1083-90. doi: 10.18632/oncotarget.1804. Oncotarget. 2014. PMID: 24658544 Free PMC article.
-
Distinct biological roles for the akt family in mammary tumor progression.Cancer Res. 2010 Jun 1;70(11):4260-4. doi: 10.1158/0008-5472.CAN-10-0266. Epub 2010 Apr 27. Cancer Res. 2010. PMID: 20424120 Free PMC article. Review.
Cited by
-
The role of immunoglobulin superfamily cell adhesion molecules in cancer metastasis.Int J Cell Biol. 2012;2012:340296. doi: 10.1155/2012/340296. Epub 2012 Jan 9. Int J Cell Biol. 2012. PMID: 22272201 Free PMC article.
-
Pparγ1 Facilitates ErbB2-Mammary Adenocarcinoma in Mice.Cancers (Basel). 2021 Apr 30;13(9):2171. doi: 10.3390/cancers13092171. Cancers (Basel). 2021. PMID: 33946495 Free PMC article.
-
Akt isoform-specific signaling in breast cancer: uncovering an anti-migratory role for palladin.Cell Adh Migr. 2011 May-Jun;5(3):211-4. doi: 10.4161/cam.5.3.15790. Epub 2011 May 1. Cell Adh Migr. 2011. PMID: 21519185 Free PMC article. Review.
-
The nuclear receptor coactivator amplified in breast cancer-1 is required for Neu (ErbB2/HER2) activation, signaling, and mammary tumorigenesis in mice.Cancer Res. 2008 May 15;68(10):3697-706. doi: 10.1158/0008-5472.CAN-07-6702. Cancer Res. 2008. PMID: 18483252 Free PMC article.
-
Tip30 deletion in MMTV-Neu mice leads to enhanced EGFR signaling and development of estrogen receptor-positive and progesterone receptor-negative mammary tumors.Cancer Res. 2010 Dec 15;70(24):10224-33. doi: 10.1158/0008-5472.CAN-10-3057. Cancer Res. 2010. PMID: 21159643 Free PMC article.
References
-
- Dudek H, Datta SR, Franke TF, Birnbaum MJ, Yao R, Cooper GM, Segal RA, Kaplan DR, Greenberg ME. Science. 1997;275:661–665. - PubMed
-
- Majewski N, Nogueira V, Bhaskar P, Coy PE, Skeen JE, Gottlob K, Chandel NS, Thompson CB, Robey RB, Hay N. Mol Cell. 2004;16:819–830. - PubMed
-
- Simpson L, Parsons R. Exp Cell Res. 2001;264:29–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA075503/CA/NCI NIH HHS/United States
- R01 CA093495/CA/NCI NIH HHS/United States
- R01CA75503/CA/NCI NIH HHS/United States
- R01 CCA093495/PHS HHS/United States
- 1P30CA56036-08/CA/NCI NIH HHS/United States
- R01 DK048910/DK/NIDDK NIH HHS/United States
- R01 CA070896/CA/NCI NIH HHS/United States
- R01 CA090764/CA/NCI NIH HHS/United States
- K01 CA085502/CA/NCI NIH HHS/United States
- R01CA107382/CA/NCI NIH HHS/United States
- 5R01DK48910/DK/NIDDK NIH HHS/United States
- R01CA70896/CA/NCI NIH HHS/United States
- R01CA090764/CA/NCI NIH HHS/United States
- K01 CA85502/CA/NCI NIH HHS/United States
- R01 CA107382/CA/NCI NIH HHS/United States
- P30 CA056036/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
