

Molecular architecture and functional model of the complete yeast ESCRT-I heterotetramer
- PMID: 17442384
- PMCID: PMC2065850
- DOI: 10.1016/j.cell.2007.03.016
Molecular architecture and functional model of the complete yeast ESCRT-I heterotetramer
Abstract
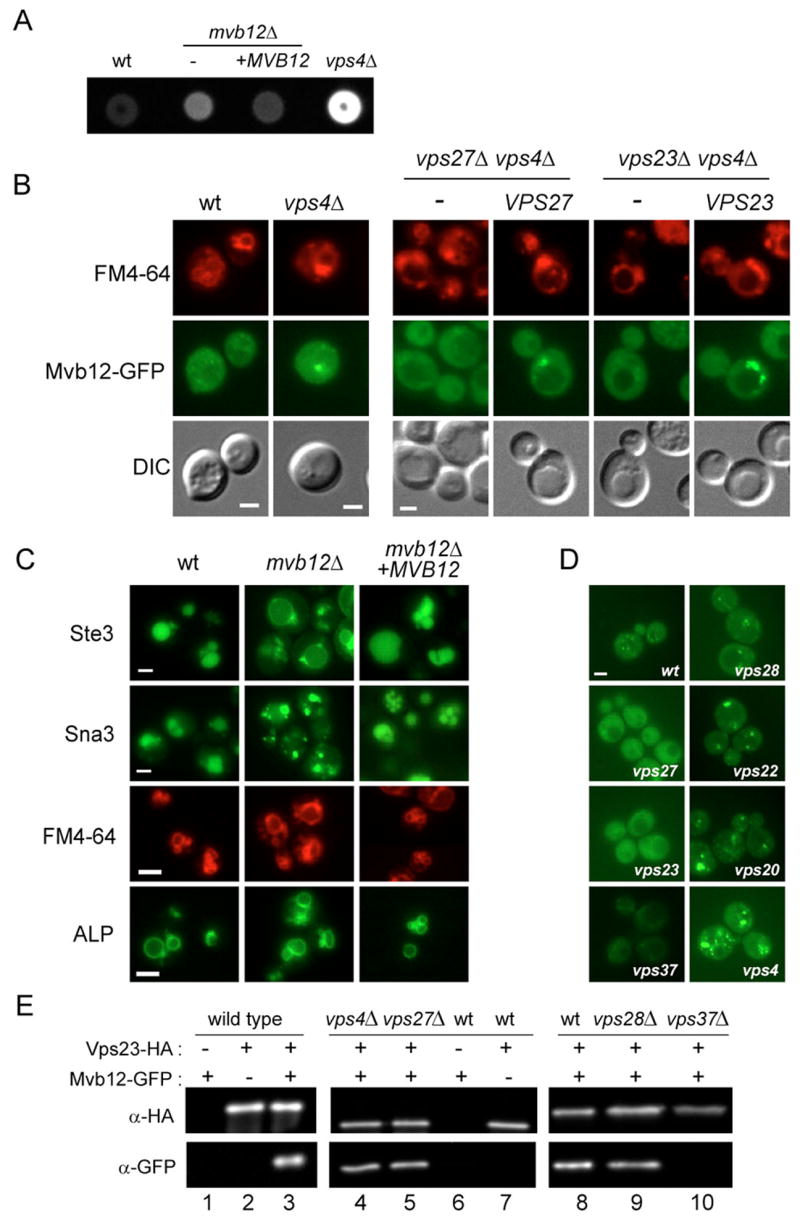
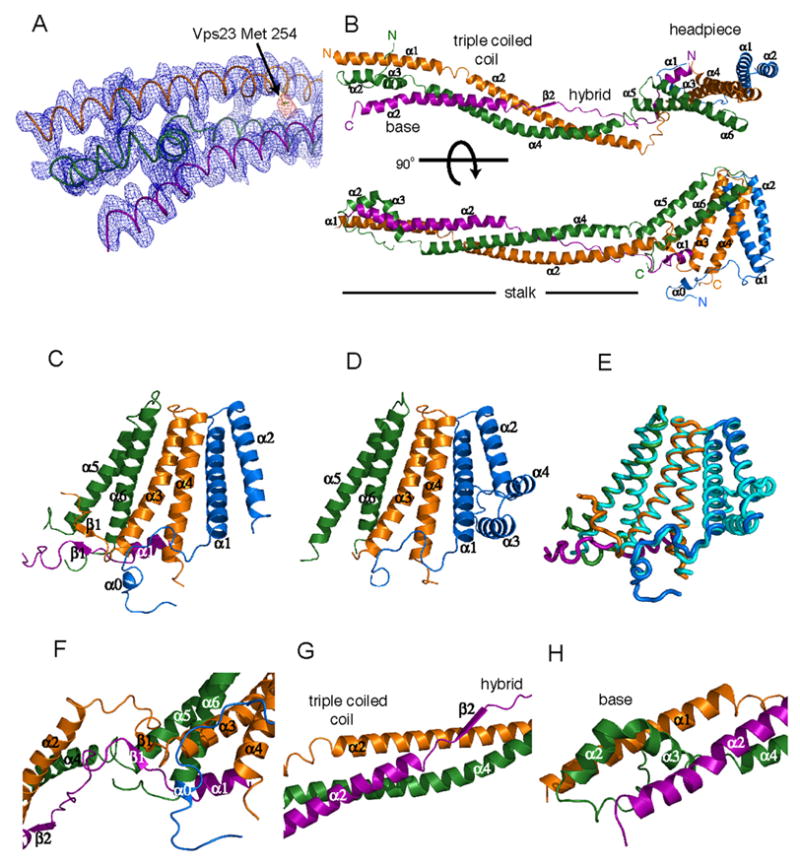
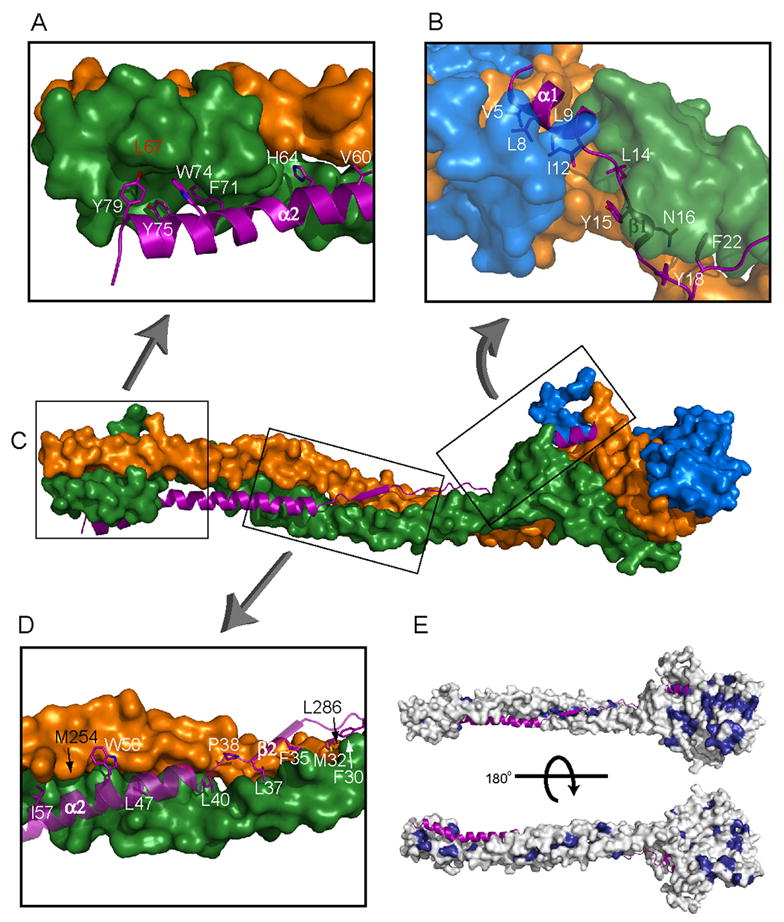
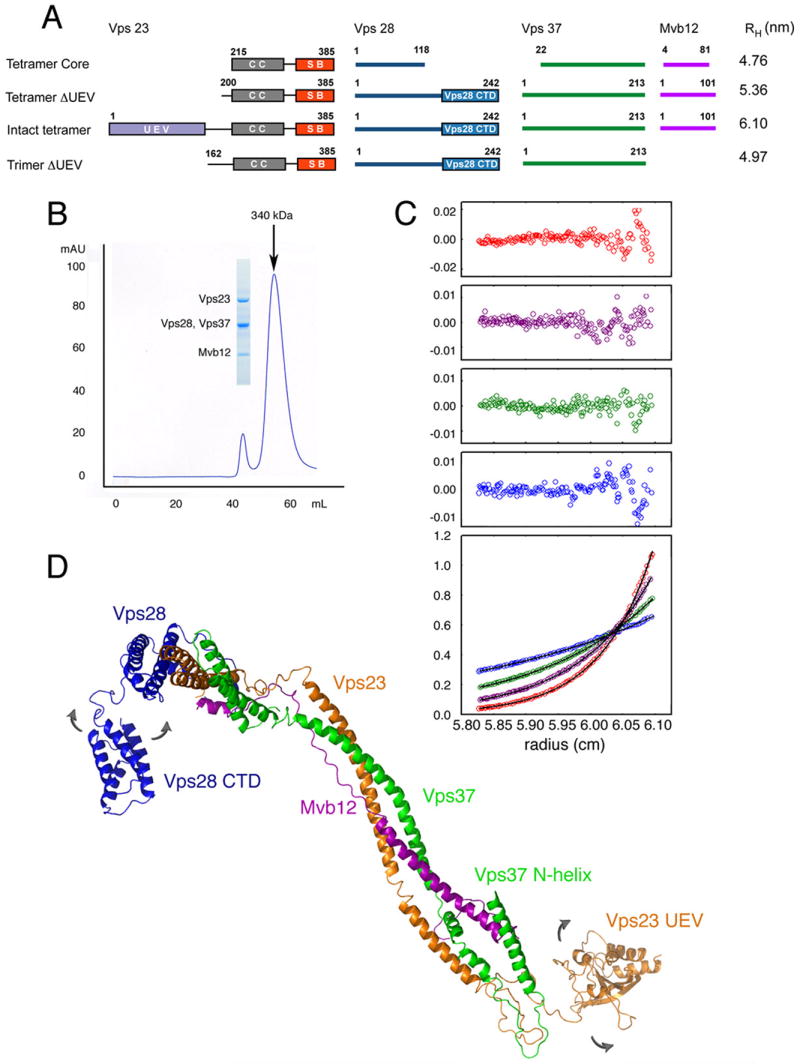
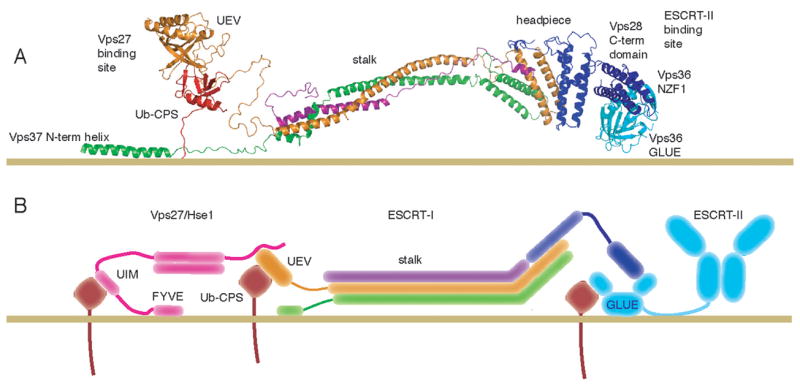
The endosomal sorting complex required for transport-I (ESCRT-I) complex, which is conserved from yeast to humans, directs the lysosomal degradation of ubiquitinated transmembrane proteins and the budding of the HIV virus. Yeast ESCRT-I contains four subunits, Vps23, Vps28, Vps37, and Mvb12. The crystal structure of the heterotetrameric ESCRT-I complex reveals a highly asymmetric complex of 1:1:1:1 subunit stoichiometry. The core complex is nearly 18 nm long and consists of a headpiece attached to a 13 nm stalk. The stalk is important for cargo sorting by ESCRT-I and is proposed to serve as a spacer regulating the correct disposition of cargo and other ESCRT components. Hydrodynamic constraints and crystallographic structures were used to generate a model of intact ESCRT-I in solution. The results show how ESCRT-I uses a combination of a rigid stalk and flexible tethers to interact with lipids, cargo, and other ESCRT complexes over a span of approximately 25 nm.
Figures
Similar articles
-
ESCRT-I core and ESCRT-II GLUE domain structures reveal role for GLUE in linking to ESCRT-I and membranes.Cell. 2006 Apr 7;125(1):99-111. doi: 10.1016/j.cell.2006.01.047. Cell. 2006. PMID: 16615893
-
Structural and functional organization of the ESCRT-I trafficking complex.Cell. 2006 Apr 7;125(1):113-26. doi: 10.1016/j.cell.2006.01.049. Cell. 2006. PMID: 16615894 Free PMC article.
-
Efficient cargo sorting by ESCRT-I and the subsequent release of ESCRT-I from multivesicular bodies requires the subunit Mvb12.Mol Biol Cell. 2007 Feb;18(2):636-45. doi: 10.1091/mbc.e06-07-0588. Epub 2006 Nov 29. Mol Biol Cell. 2007. PMID: 17135292 Free PMC article.
-
The emerging shape of the ESCRT machinery.Nat Rev Mol Cell Biol. 2007 May;8(5):355-68. doi: 10.1038/nrm2162. Nat Rev Mol Cell Biol. 2007. PMID: 17450176 Review.
-
Structures, Functions, and Dynamics of ESCRT-III/Vps4 Membrane Remodeling and Fission Complexes.Annu Rev Cell Dev Biol. 2018 Oct 6;34:85-109. doi: 10.1146/annurev-cellbio-100616-060600. Epub 2018 Aug 10. Annu Rev Cell Dev Biol. 2018. PMID: 30095293 Free PMC article. Review.
Cited by
-
Wrapping up the bad news: HIV assembly and release.Retrovirology. 2013 Jan 10;10:5. doi: 10.1186/1742-4690-10-5. Retrovirology. 2013. PMID: 23305486 Free PMC article. Review.
-
SProtP: a web server to recognize those short-lived proteins based on sequence-derived features in human cells.PLoS One. 2011;6(11):e27836. doi: 10.1371/journal.pone.0027836. Epub 2011 Nov 16. PLoS One. 2011. PMID: 22114707 Free PMC article.
-
Phosphoinositides in the mammalian endo-lysosomal network.Subcell Biochem. 2012;59:65-110. doi: 10.1007/978-94-007-3015-1_3. Subcell Biochem. 2012. PMID: 22374088 Free PMC article. Review.
-
ESCRT machinery mediates selective microautophagy of endoplasmic reticulum in yeast.EMBO J. 2020 Jan 15;39(2):e102586. doi: 10.15252/embj.2019102586. Epub 2019 Dec 5. EMBO J. 2020. PMID: 31802527 Free PMC article.
-
No strings attached: the ESCRT machinery in viral budding and cytokinesis.J Cell Sci. 2009 Jul 1;122(Pt 13):2167-77. doi: 10.1242/jcs.028308. J Cell Sci. 2009. PMID: 19535732 Free PMC article.
References
-
- Babst M. A Protein’s Final ESCRT. Traffic. 2005;6:2–9. - PubMed
-
- Bilodeau PS, Urbanowski JL, Winistorfer SC, Piper RC. The Vps27p-Hse1p complex binds ubiquitin and mediates endosomal protein sorting. Nat Cell Biol. 2002;4:534–539. - PubMed
-
- Bowers K, Stevens TH. Protein transport from the late Golgi to the vacuole in the yeast Saccharomyces cerevisiae. Biochimica Et Biophysica Acta-Molecular Cell Research. 2005;1744:438–454. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
