

Activation of endosomal dynein motors by stepwise assembly of Rab7-RILP-p150Glued, ORP1L, and the receptor betalll spectrin
- PMID: 17283181
- PMCID: PMC2063981
- DOI: 10.1083/jcb.200606077
Activation of endosomal dynein motors by stepwise assembly of Rab7-RILP-p150Glued, ORP1L, and the receptor betalll spectrin
Abstract
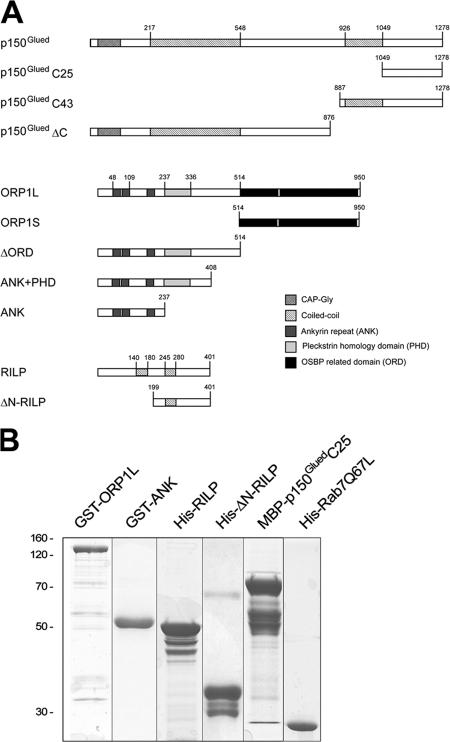
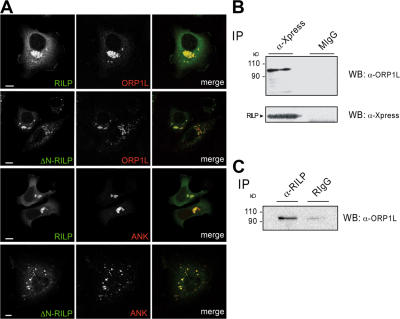
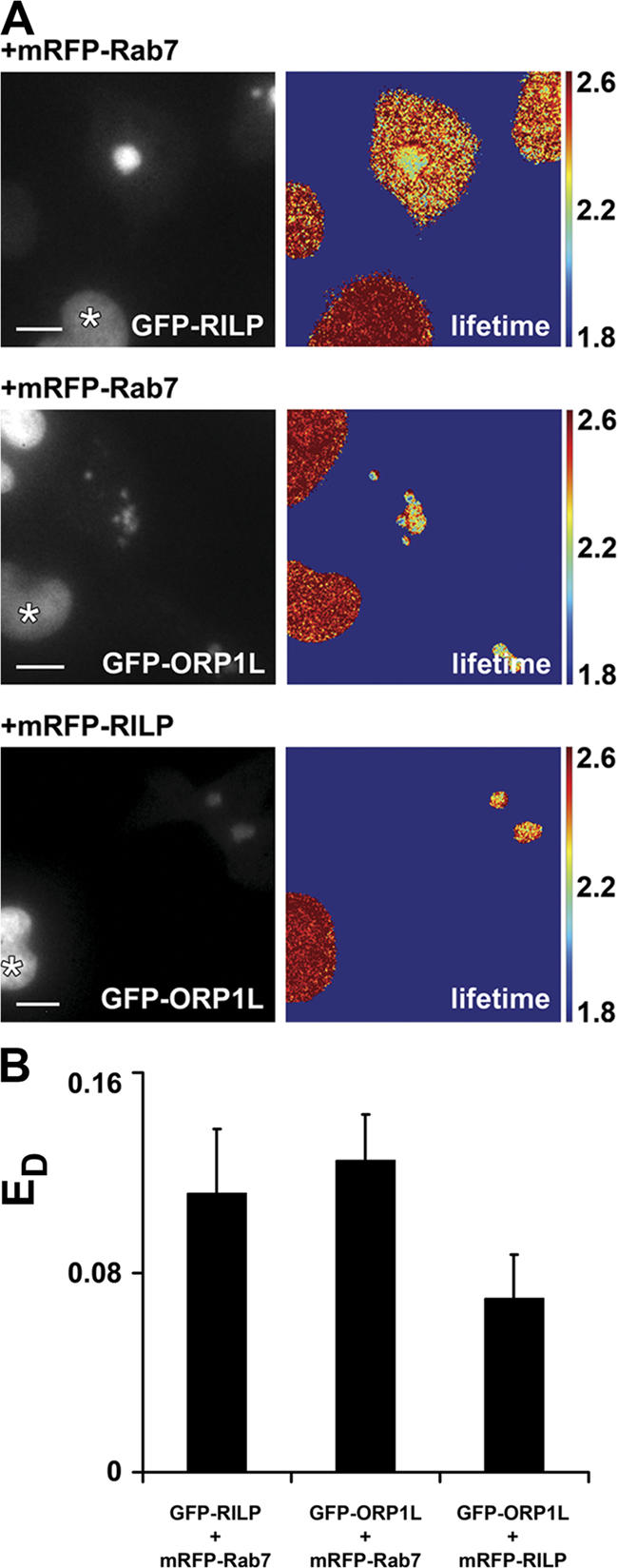
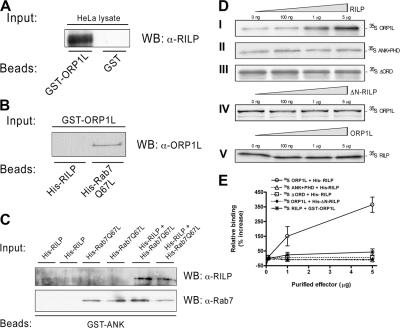
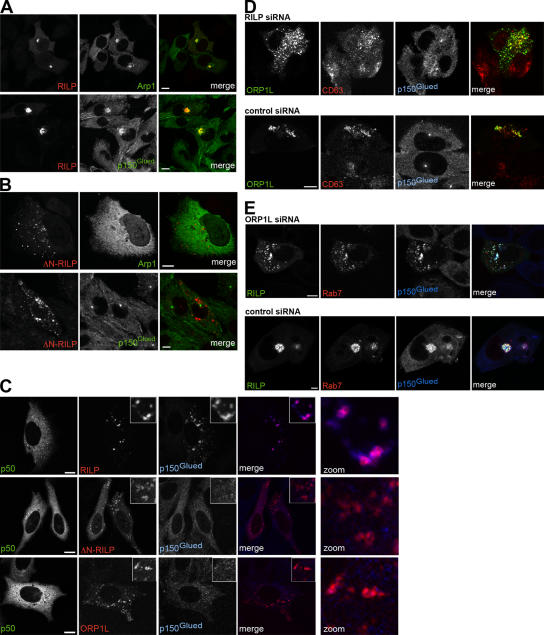
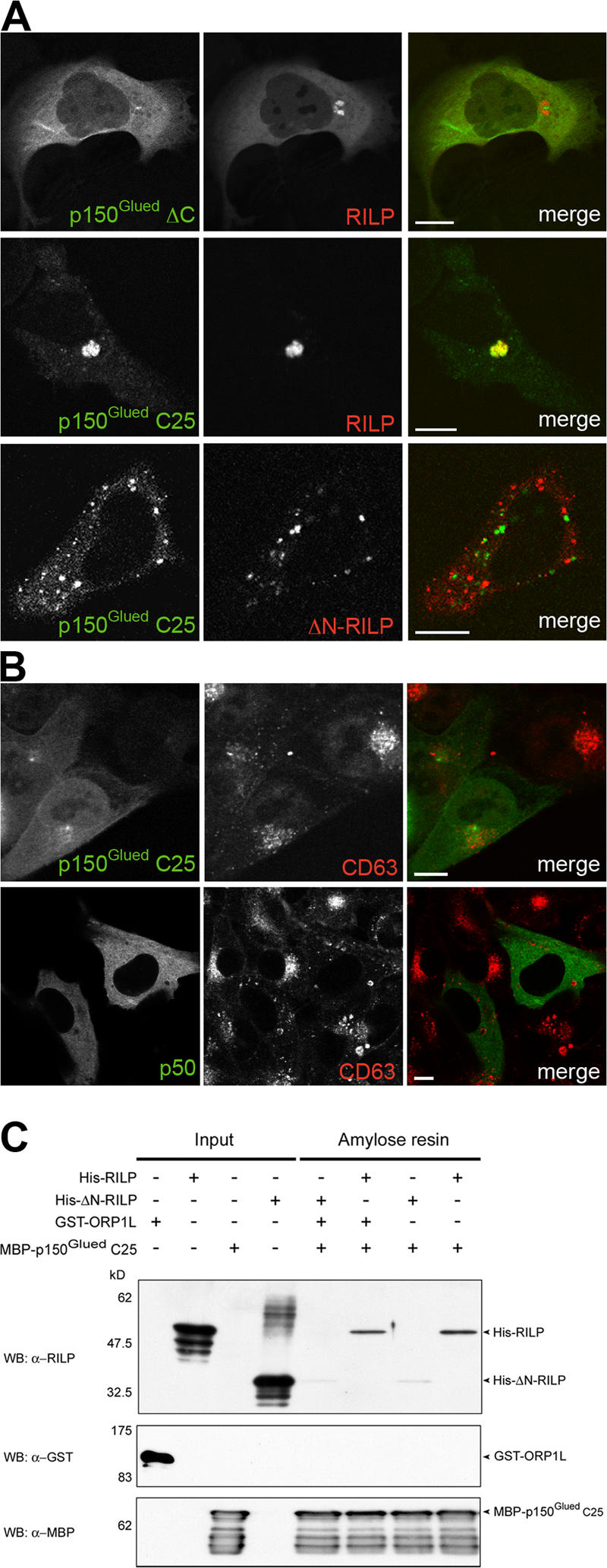
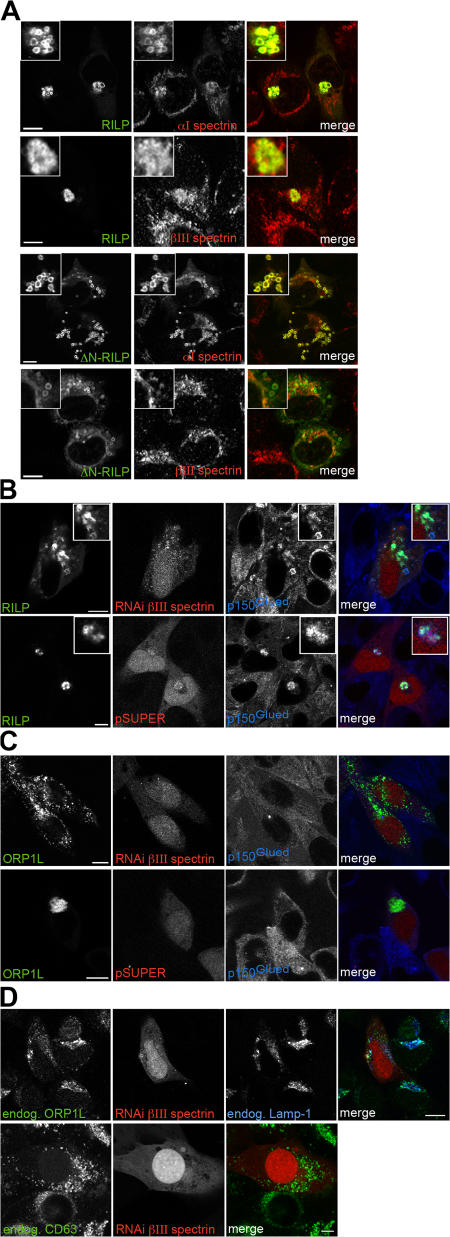
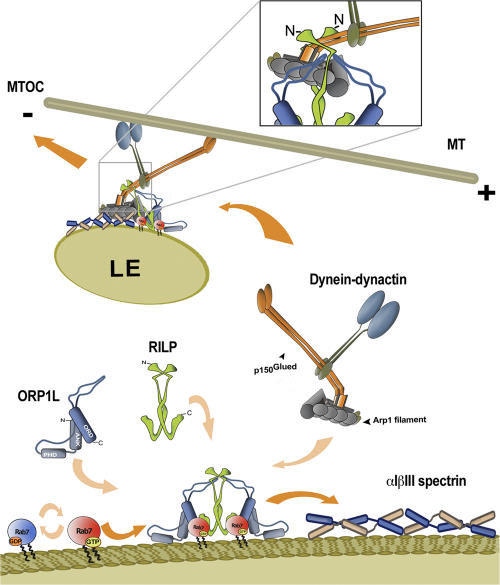
The small GTPase Rab7 controls late endocytic transport by the minus end-directed motor protein complex dynein-dynactin, but how it does this is unclear. Rab7-interacting lysosomal protein (RILP) and oxysterol-binding protein-related protein 1L (ORP1L) are two effectors of Rab7. We show that GTP-bound Rab7 simultaneously binds RILP and ORP1L to form a RILP-Rab7-ORP1L complex. RILP interacts directly with the C-terminal 25-kD region of the dynactin projecting arm p150(Glued), which is required for dynein motor recruitment to late endocytic compartments (LEs). Still, p150(Glued) recruitment by Rab7-RILP does not suffice to induce dynein-driven minus-end transport of LEs. ORP1L, as well as betaIII spectrin, which is the general receptor for dynactin on vesicles, are essential for dynein motor activity. Our results illustrate that the assembly of microtubule motors on endosomes involves a cascade of linked events. First, Rab7 recruits two effectors, RILP and ORP1L, to form a tripartite complex. Next, RILP directly binds to the p150(Glued) dynactin subunit to recruit the dynein motor. Finally, the specific dynein motor receptor Rab7-RILP is transferred by ORP1L to betaIII spectrin. Dynein will initiate translocation of late endosomes to microtubule minus ends only after interacting with betaIII spectrin, which requires the activities of Rab7-RILP and ORP1L.
Figures
Similar articles
-
Cholesterol sensor ORP1L contacts the ER protein VAP to control Rab7-RILP-p150 Glued and late endosome positioning.J Cell Biol. 2009 Jun 29;185(7):1209-25. doi: 10.1083/jcb.200811005. J Cell Biol. 2009. PMID: 19564404 Free PMC article.
-
Late endosomal transport and tethering are coupled processes controlled by RILP and the cholesterol sensor ORP1L.J Cell Sci. 2013 Aug 1;126(Pt 15):3462-74. doi: 10.1242/jcs.129270. Epub 2013 May 31. J Cell Sci. 2013. PMID: 23729732
-
The Rab7 effector protein RILP controls lysosomal transport by inducing the recruitment of dynein-dynactin motors.Curr Biol. 2001 Oct 30;11(21):1680-5. doi: 10.1016/s0960-9822(01)00531-0. Curr Biol. 2001. PMID: 11696325
-
Rab7: role of its protein interaction cascades in endo-lysosomal traffic.Cell Signal. 2011 Mar;23(3):516-21. doi: 10.1016/j.cellsig.2010.09.012. Epub 2010 Sep 21. Cell Signal. 2011. PMID: 20851765 Review.
-
Dynein motility: four heads are better than two.Curr Biol. 2005 Dec 6;15(23):R970-2. doi: 10.1016/j.cub.2005.11.021. Curr Biol. 2005. PMID: 16332531 Review.
Cited by
-
Sterol-dependent nuclear import of ORP1S promotes LXR regulated trans-activation of apoE.Exp Cell Res. 2012 Oct 1;318(16):2128-42. doi: 10.1016/j.yexcr.2012.06.012. Epub 2012 Jun 20. Exp Cell Res. 2012. PMID: 22728266 Free PMC article.
-
Interactions between the Coxiella burnetii parasitophorous vacuole and the endoplasmic reticulum involve the host protein ORP1L.Cell Microbiol. 2017 Jan;19(1):10.1111/cmi.12637. doi: 10.1111/cmi.12637. Epub 2016 Jul 15. Cell Microbiol. 2017. PMID: 27345457 Free PMC article.
-
HkRP3 is a microtubule-binding protein regulating lytic granule clustering and NK cell killing.J Immunol. 2015 Apr 15;194(8):3984-96. doi: 10.4049/jimmunol.1402897. Epub 2015 Mar 11. J Immunol. 2015. PMID: 25762780 Free PMC article.
-
Structure-function analyses of the small GTPase Rab35 and its effector protein centaurin-β2/ACAP2 during neurite outgrowth of PC12 cells.J Biol Chem. 2015 Apr 3;290(14):9064-74. doi: 10.1074/jbc.M114.611301. Epub 2015 Feb 18. J Biol Chem. 2015. PMID: 25694427 Free PMC article.
-
Genome-scale requirements for dynein-based trafficking revealed by a high-content arrayed CRISPR screen.bioRxiv [Preprint]. 2023 Mar 1:2023.03.01.530592. doi: 10.1101/2023.03.01.530592. bioRxiv. 2023. Update in: J Cell Biol. 2024 May 6;223(5):e202306048. doi: 10.1083/jcb.202306048. PMID: 36909483 Free PMC article. Updated. Preprint.
References
-
- Bastiaens, P.I., and A. Squire. 1999. Fluorescence lifetime imaging microscopy: spatial resolution of biochemical processes in the cell. Trends Cell Biol. 9:48–52. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
