

Impaired RNA incorporation and dimerization in live attenuated leader-variants of SIVmac239
- PMID: 17184529
- PMCID: PMC1766366
- DOI: 10.1186/1742-4690-3-96
Impaired RNA incorporation and dimerization in live attenuated leader-variants of SIVmac239
Abstract
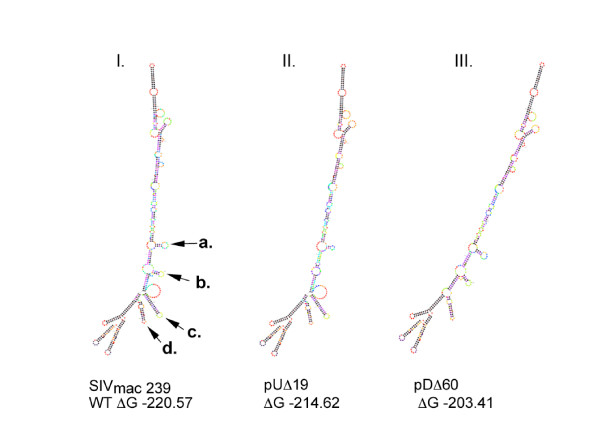
Background: The 5' untranslated region (UTR) or leader sequence of simian immunodeficiency virus (SIVmac239) is multifunctional and harbors the regulatory elements for viral replication, persistence, gene translation, expression, and the packaging and dimerization of viral genomic RNA (vRNA). We have constructed a series of deletions in the SIVmac239 leader sequence in order to determine the involvement of this region in both the packaging and dimerization of viral genomic RNA. We also assessed the impact of these deletions upon viral infectiousness, replication kinetics and gene expression in cell lines and monkey peripheral blood mononuclear cells (PBMC).
Results: Regions on both sides of the major splice donor (SD) were found to be necessary for the efficiency and specificity of viral genome packaging. However, stem-loop1 is critical for both RNA encapsidation and dimerization. Downstream elements between the splice donor and the initiation site of SIV-Gag have additive effects on RNA packaging and contribute to a lesser degree to RNA dimerization. The targeted disruption of structures on both sides of the SD also severely impacts viral infectiousness, gene expression and replication in both CEMx174 cells and rhesus PBMC.
Conclusion: In the leader region of SIVmac239, stem-loop1 functions as the primary determinant for both RNA encapsidation and dimerization. Downstream elements between the splice donor and the translational initiation site of SIV-Gag are classified as secondary determinants and play a role in dimerization. Collectively, these data signify a linkage between the primary encapsidation determinant of SIVmac239 and RNA dimerization.
Figures


Similar articles
-
Novel, live attenuated simian immunodeficiency virus constructs containing major deletions in leader RNA sequences.J Virol. 2001 Mar;75(6):2776-85. doi: 10.1128/JVI.75.6.2776-2785.2001. J Virol. 2001. PMID: 11222701 Free PMC article.
-
Sequences in the 5' leader of Mason-Pfizer monkey virus which affect viral particle production and genomic RNA packaging: development of MPMV packaging cell lines.Virology. 2001 Sep 15;288(1):81-8. doi: 10.1006/viro.2001.1061. Virology. 2001. PMID: 11543660
-
Mutational analysis of the 5' leader region of simian foamy virus type 1.Virology. 2000 Aug 15;274(1):203-12. doi: 10.1006/viro.2000.0423. Virology. 2000. PMID: 10936101
-
Flavors of Flaviviral RNA Structure: towards an Integrated View of RNA Function from Translation through Encapsidation.Bioessays. 2019 Aug;41(8):e1900003. doi: 10.1002/bies.201900003. Epub 2019 Jun 18. Bioessays. 2019. PMID: 31210384 Free PMC article. Review.
-
Structural determinants and mechanism of HIV-1 genome packaging.J Mol Biol. 2011 Jul 22;410(4):609-33. doi: 10.1016/j.jmb.2011.04.029. J Mol Biol. 2011. PMID: 21762803 Free PMC article. Review.
Cited by
-
Sequences within both the 5' UTR and Gag are required for optimal in vivo packaging and propagation of mouse mammary tumor virus (MMTV) genomic RNA.PLoS One. 2012;7(10):e47088. doi: 10.1371/journal.pone.0047088. Epub 2012 Oct 16. PLoS One. 2012. PMID: 23077548 Free PMC article.
-
HIV-2 genome dimerization is required for the correct processing of Gag: a second-site reversion in matrix can restore both processes in dimerization-impaired mutant viruses.J Virol. 2012 May;86(10):5867-76. doi: 10.1128/JVI.00124-12. Epub 2012 Mar 14. J Virol. 2012. PMID: 22419802 Free PMC article.
-
Retroviral RNA Dimerization: From Structure to Functions.Front Microbiol. 2018 Mar 22;9:527. doi: 10.3389/fmicb.2018.00527. eCollection 2018. Front Microbiol. 2018. PMID: 29623074 Free PMC article. Review.
-
Cross- and Co-Packaging of Retroviral RNAs and Their Consequences.Viruses. 2016 Oct 11;8(10):276. doi: 10.3390/v8100276. Viruses. 2016. PMID: 27727192 Free PMC article. Review.
-
HIV-2 RNA dimerization is regulated by intramolecular interactions in vitro.RNA. 2007 Aug;13(8):1341-54. doi: 10.1261/rna.483807. Epub 2007 Jun 25. RNA. 2007. PMID: 17592043 Free PMC article.
References
-
- Rein A. Retroviral RNA packaging: a review. Arch Virol Suppl. 1994;9:513–522. - PubMed
-
- Russell RS, Hu J, Beriault V, Mouland AJ, Laughrea M, Kleiman L, Wainberg MA, Liang C. Sequences downstream of the 5' splice donor site are required for both packaging and dimerization of human immunodeficiency virus type 1 RNA. J Virol. 2003;77:84–96. doi: 10.1128/JVI.77.1.84-96.2003. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
