

Phosphorylation-dependent ubiquitination of cyclin D1 by the SCF(FBX4-alphaB crystallin) complex
- PMID: 17081987
- PMCID: PMC1702390
- DOI: 10.1016/j.molcel.2006.09.007
Phosphorylation-dependent ubiquitination of cyclin D1 by the SCF(FBX4-alphaB crystallin) complex
Abstract
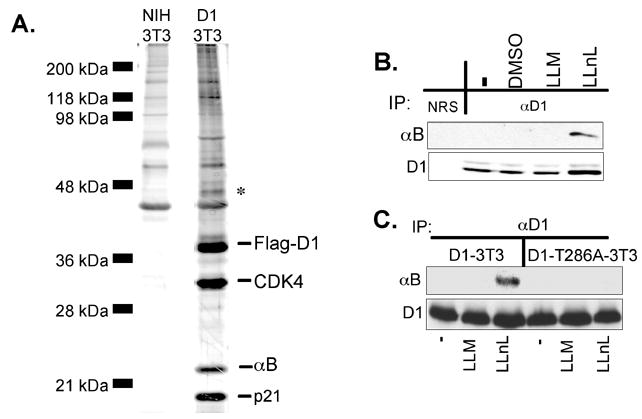
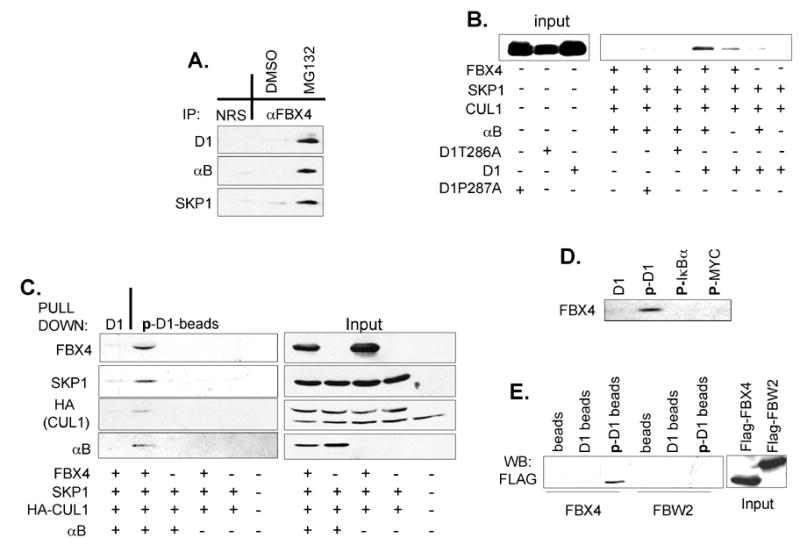
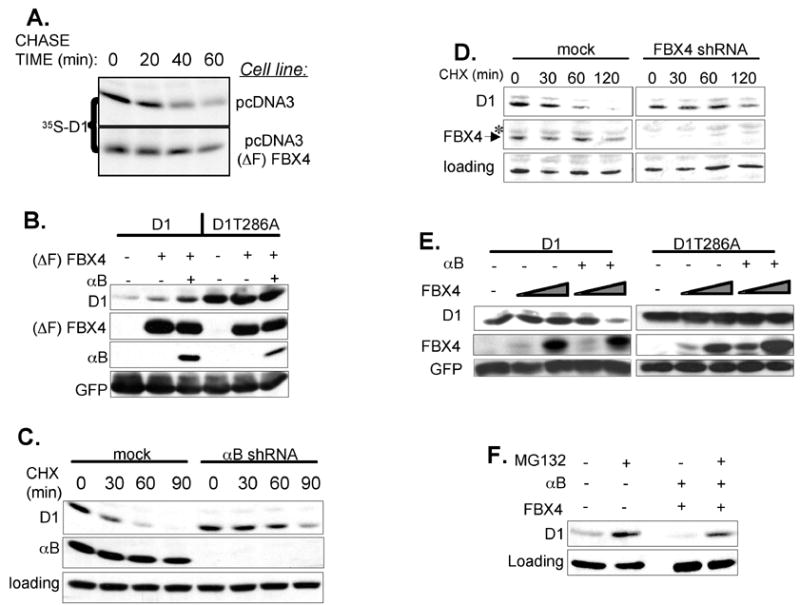
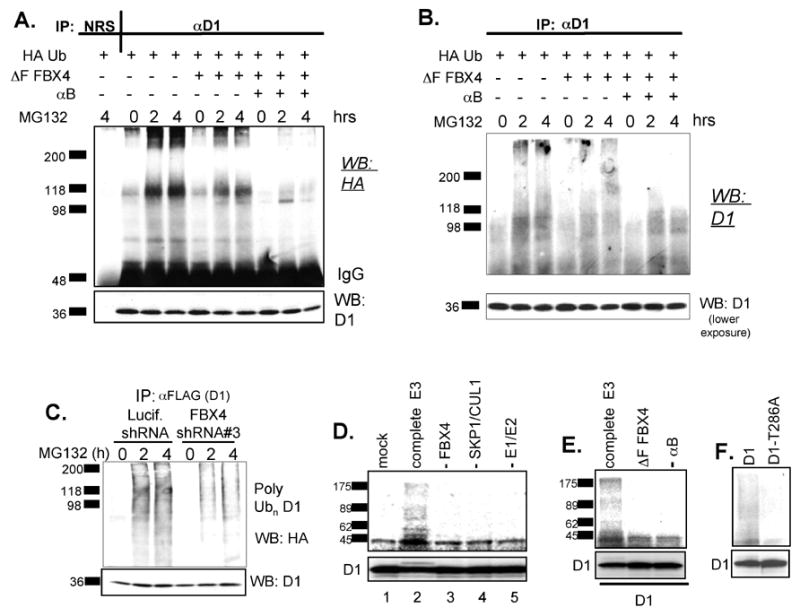
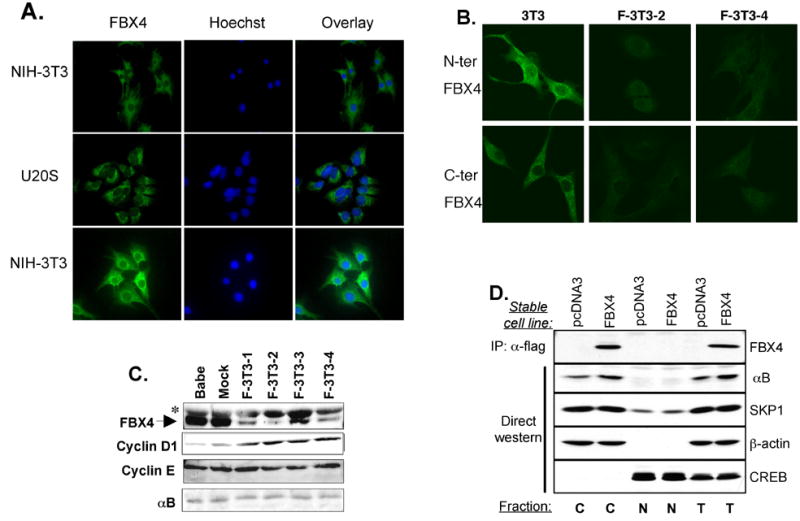
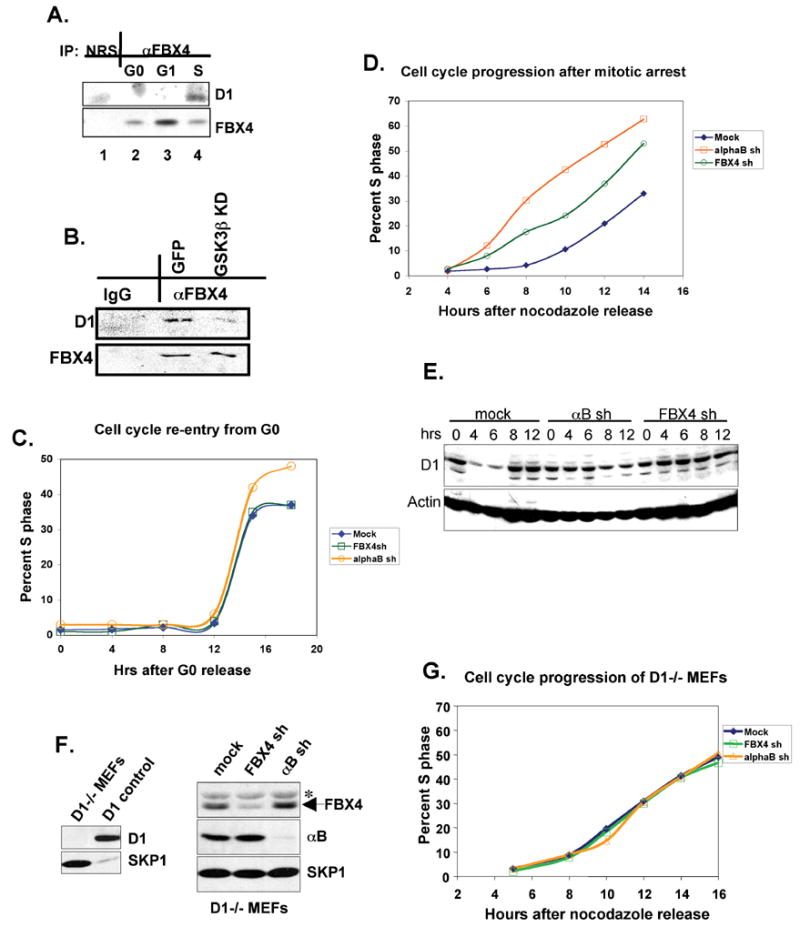
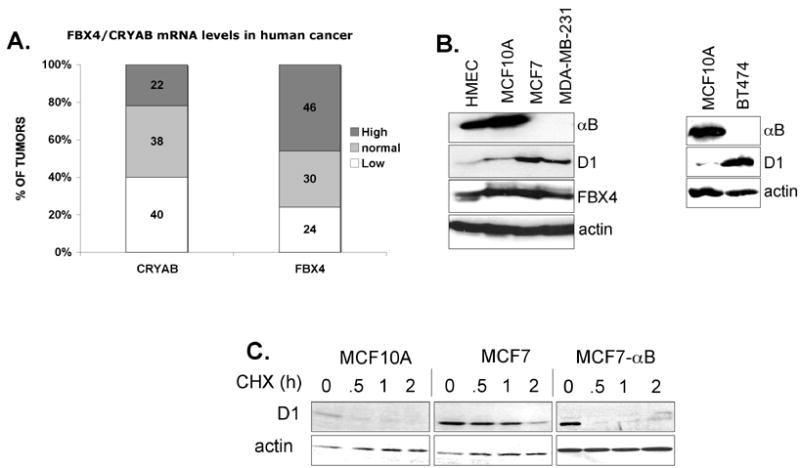
Growth factor-dependent accumulation of the cyclin D1 proto-oncogene is balanced by its rapid phosphorylation-dependent proteolysis. Degradation is triggered by threonine 286 phosphorylation, which promotes its ubiquitination by an unknown E3 ligase. We demonstrate that Thr286-phosphorylated cyclin D1 is recognized by a Skp1-Cul1-F box (SCF) ubiquitin ligase where FBX4 and alphaB crystallin govern substrate specificity. Overexpression of FBX4 and alphaB crystallin triggered cyclin D1 ubiquitination and increased cyclin D1 turnover. Impairment of SCF(FBX4-alphaB crystallin) function attenuated cyclin D1 ubiquitination, promoting cyclin D1 overexpression and accelerated cell-cycle progression. Purified SCF(FBX4-alphaB crystallin) catalyzed polyubiquitination of cyclin D1 in vitro. Consistent with a putative role for a cyclin D1 E3 ligase in tumorigenesis, FBX4 and alphaB crystallin expression was reduced in tumor-derived cell lines and a subset of primary human cancers that overexpress cyclin D1. We conclude that SCF(FBX4-alphaB crystallin) is an E3 ubiquitin ligase that promotes ubiquitin-dependent degradation of Thr286-phosphorylated cyclin D1.
Figures
Similar articles
-
SCF(Fbx4/alphaB-crystallin) E3 ligase: when one is not enough.Cell Cycle. 2008 Oct;7(19):2983-6. doi: 10.4161/cc.7.19.6775. Epub 2008 Oct 12. Cell Cycle. 2008. PMID: 18818515 Review.
-
Lysine 269 is essential for cyclin D1 ubiquitylation by the SCF(Fbx4/alphaB-crystallin) ligase and subsequent proteasome-dependent degradation.Oncogene. 2009 Dec 10;28(49):4317-25. doi: 10.1038/onc.2009.287. Oncogene. 2009. PMID: 19767775 Free PMC article.
-
Phosphorylation-dependent regulation of SCF(Fbx4) dimerization and activity involves a novel component, 14-3-3ɛ.Oncogene. 2011 Apr 28;30(17):1995-2002. doi: 10.1038/onc.2010.584. Epub 2011 Jan 17. Oncogene. 2011. PMID: 21242966 Free PMC article.
-
The small heat-shock protein alpha B-crystallin promotes FBX4-dependent ubiquitination.J Biol Chem. 2003 Feb 14;278(7):4699-704. doi: 10.1074/jbc.M211403200. Epub 2002 Dec 4. J Biol Chem. 2003. PMID: 12468532
-
The Biology of F-box Proteins: The SCF Family of E3 Ubiquitin Ligases.Adv Exp Med Biol. 2020;1217:111-122. doi: 10.1007/978-981-15-1025-0_8. Adv Exp Med Biol. 2020. PMID: 31898225 Review.
Cited by
-
Mechanism-based screen for G1/S checkpoint activators identifies a selective activator of EIF2AK3/PERK signalling.PLoS One. 2012;7(1):e28568. doi: 10.1371/journal.pone.0028568. Epub 2012 Jan 12. PLoS One. 2012. PMID: 22253692 Free PMC article.
-
Non-Hsp genes are essential for HSF1-mediated maintenance of whole body homeostasis.Exp Anim. 2015;64(4):397-406. doi: 10.1538/expanim.15-0023. Epub 2015 Aug 4. Exp Anim. 2015. PMID: 26251235 Free PMC article.
-
Small heat shock proteins target mutant cystic fibrosis transmembrane conductance regulator for degradation via a small ubiquitin-like modifier-dependent pathway.Mol Biol Cell. 2013 Jan;24(2):74-84. doi: 10.1091/mbc.E12-09-0678. Epub 2012 Nov 14. Mol Biol Cell. 2013. PMID: 23155000 Free PMC article.
-
Promoter hypermethylation of FBXO32, a novel TGF-beta/SMAD4 target gene and tumor suppressor, is associated with poor prognosis in human ovarian cancer.Lab Invest. 2010 Mar;90(3):414-25. doi: 10.1038/labinvest.2009.138. Epub 2010 Jan 11. Lab Invest. 2010. PMID: 20065949 Free PMC article.
-
The cell biology of the unfolded protein response.Gastroenterology. 2011 Jul;141(1):38-41, 41.e1-2. doi: 10.1053/j.gastro.2011.05.018. Epub 2011 May 24. Gastroenterology. 2011. PMID: 21620842 Free PMC article. Review.
References
-
- Alt JR, Gladden AB, Diehl JA. p21(Cip1) Promotes cyclin D1 nuclear accumulation via direct inhibition of nuclear export. J Biol Chem. 2002;277:8517–8523. - PubMed
-
- Bai F, Xi JH, Wawrousek EF, Fleming TP, Andley UP. Hyperproliferation and p53 status of lens epithelial cells derived from alphaB-crystallin knockout mice. J Biol Chem. 2003;278:36876–36886. - PubMed
-
- Bashir T, Dorrello NV, Amador V, Guardavaccaro D, Pagano M. Control of the SCF(Skp2-Cks1) ubiquitin ligase by the APC/C(Cdh1) ubiquitin ligase. Nature. 2004;428:190–193. - PubMed
-
- Bax B, Carter PS, Lewis C, Guy AR, Bridges A, Tanner R, Pettman G, Mannix C, Culbert AA, Brown MJ, et al. The structure of phosphorylated GSK-3beta complexed with a peptide, FRATtide, that inhibits beta-catenin phosphorylation. Structure (Camb) 2001;9:1143–1152. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
