

The multifunctional beta-oxidation enzyme is required for full symptom development by the biotrophic maize pathogen Ustilago maydis
- PMID: 16998075
- PMCID: PMC1694828
- DOI: 10.1128/EC.00231-06
The multifunctional beta-oxidation enzyme is required for full symptom development by the biotrophic maize pathogen Ustilago maydis
Abstract
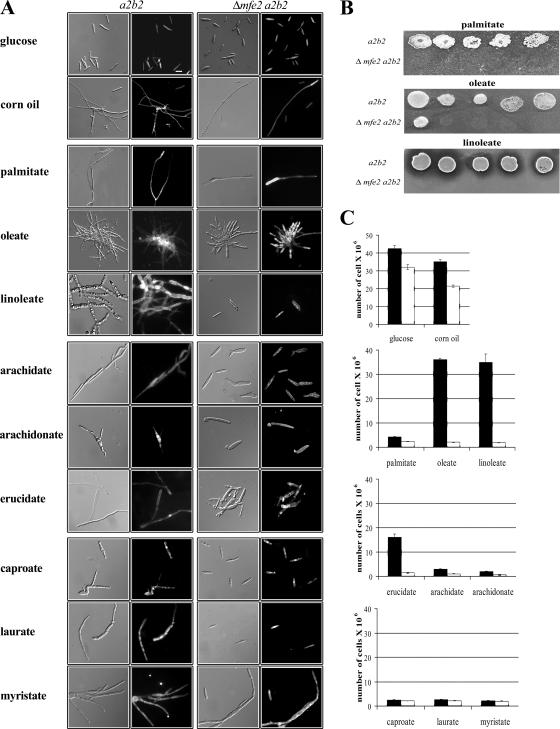
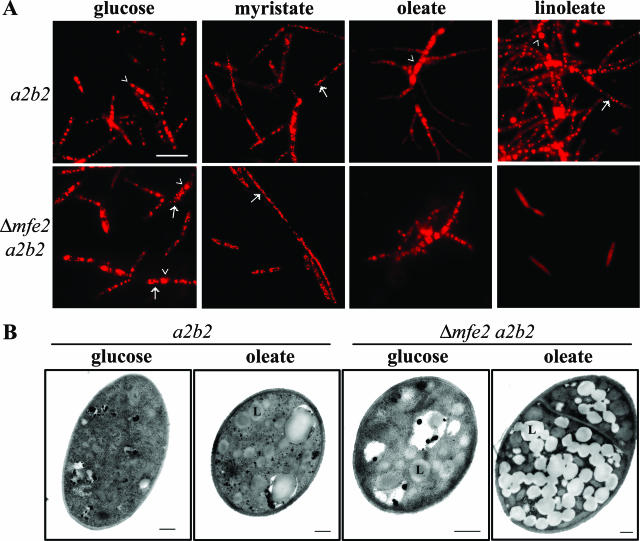
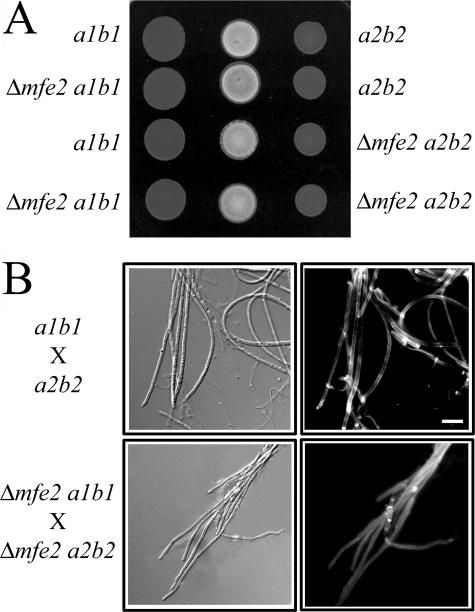
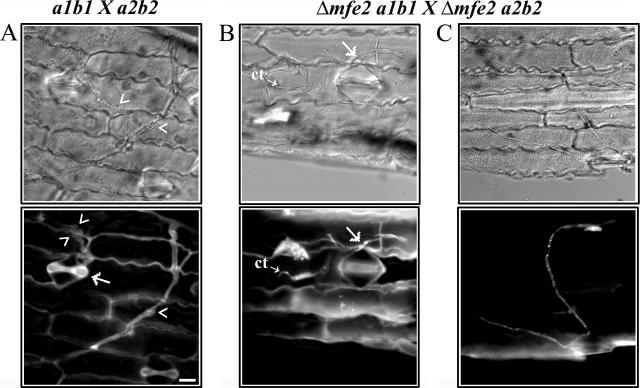
The transition from yeast-like to filamentous growth in the biotrophic fungal phytopathogen Ustilago maydis is a crucial event for pathogenesis. Previously, we showed that fatty acids induce filamentation in U. maydis and that the resulting hyphal cells resemble the infectious filaments observed in planta. To explore the potential metabolic role of lipids in the morphological transition and in pathogenic development in host tissue, we deleted the mfe2 gene encoding the multifunctional enzyme that catalyzes the second and third reactions in beta-oxidation of fatty acids in peroxisomes. The growth of the strains defective in mfe2 was attenuated on long-chain fatty acids and abolished on very-long-chain fatty acids. The mfe2 gene was not generally required for the production of filaments during mating in vitro, but loss of the gene blocked extensive proliferation of fungal filaments in planta. Consistent with this observation, mfe2 mutants exhibited significantly reduced virulence in that only 27% of infected seedlings produced tumors compared to 88% tumor production upon infection by wild-type strains. Similarly, a defect in virulence was observed in developing ears upon infection of mature maize plants. Specifically, the absence of the mfe2 gene delayed the development of teliospores within mature tumor tissue. Overall, these results indicate that the ability to utilize host lipids contributes to the pathogenic development of U. maydis.
Figures

Similar articles
-
Defects in mitochondrial and peroxisomal β-oxidation influence virulence in the maize pathogen Ustilago maydis.Eukaryot Cell. 2012 Aug;11(8):1055-66. doi: 10.1128/EC.00129-12. Epub 2012 Jun 15. Eukaryot Cell. 2012. PMID: 22707484 Free PMC article.
-
An Ustilago maydis septin is required for filamentous growth in culture and for full symptom development on maize.Eukaryot Cell. 2005 Dec;4(12):2044-56. doi: 10.1128/EC.4.12.2044-2056.2005. Eukaryot Cell. 2005. PMID: 16339722 Free PMC article.
-
Two members of the Ustilago maydis velvet family influence teliospore development and virulence on maize seedlings.Fungal Genet Biol. 2013 Dec;61:111-9. doi: 10.1016/j.fgb.2013.09.002. Epub 2013 Sep 21. Fungal Genet Biol. 2013. PMID: 24064149
-
Control of mating and development in Ustilago maydis.Antonie Van Leeuwenhoek. 1994;65(3):191-7. doi: 10.1007/BF00871946. Antonie Van Leeuwenhoek. 1994. PMID: 7847885 Review.
-
The secretome of the maize pathogen Ustilago maydis.Fungal Genet Biol. 2008 Aug;45 Suppl 1:S63-70. doi: 10.1016/j.fgb.2008.03.012. Epub 2008 Mar 31. Fungal Genet Biol. 2008. PMID: 18456523 Review.
Cited by
-
The cAMP/Protein Kinase A Pathway and Virulence in Cryptococcus neoformans.Mycobiology. 2011 Sep;39(3):143-50. doi: 10.5941/MYCO.2011.39.3.143. Epub 2011 Sep 27. Mycobiology. 2011. PMID: 22783095 Free PMC article.
-
Peroxisomal and mitochondrial β-oxidation pathways influence the virulence of the pathogenic fungus Cryptococcus neoformans.Eukaryot Cell. 2012 Aug;11(8):1042-54. doi: 10.1128/EC.00128-12. Epub 2012 Jun 15. Eukaryot Cell. 2012. PMID: 22707485 Free PMC article.
-
The putative phospholipase Lip2 counteracts oxidative damage and influences the virulence of Ustilago maydis.Mol Plant Pathol. 2017 Feb;18(2):210-221. doi: 10.1111/mpp.12391. Epub 2016 Apr 21. Mol Plant Pathol. 2017. PMID: 26950180 Free PMC article.
-
Identification and molecular detection of the pathogen of Phalaenopsis leaf yellowing through genome analysis.Front Microbiol. 2024 Sep 24;15:1431813. doi: 10.3389/fmicb.2024.1431813. eCollection 2024. Front Microbiol. 2024. PMID: 39403082 Free PMC article.
-
The AP-1-like transcription factor ChAP1 balances tolerance and cell death in the response of the maize pathogen Cochliobolus heterostrophus to a plant phenolic.Curr Genet. 2020 Feb;66(1):187-203. doi: 10.1007/s00294-019-01012-7. Epub 2019 Jul 16. Curr Genet. 2020. PMID: 31312934
References
-
- Andrews, D. L., J. D. Egan, M. E. Mayorga, and S. E. Gold. 2000. The Ustilago maydis ubc4 and ubc5 genes encode members of a MAP kinase cascade required for filamentous growth. Mol. Plant-Microbe Interact. 13:781-786. - PubMed
-
- Banuett, F., and I. Herskowitz. 1996. Discrete developmental stages during teliospore formation in the corn smut fungus, Ustilago maydis. Development 122:2965-2976. - PubMed
-
- Banuett, F., and I. Herskowitz. 1994. Identification of fuz7, a Ustilago maydis MEK/MAPKK homolog required for a-locus-dependent and -independent steps in the fungal life cycle. Genes Dev. 8:1367-1378. - PubMed
-
- Barbosa, A. C., A. E. Carmo, L. Graf, R. Tomaz, C. F. Souza, J. Mendes, M. A. Randi, D. Buchi, and R. J. Schadeck. 2006. Morphology and lipid body and vacuole dynamics during secondary conidia formation in Colletotrichum acutatum: laser scanning confocal analysis. Can. J. Microbiol. 52:117-124. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
