

Betaherpesvirus-conserved cytomegalovirus tegument protein ppUL32 (pp150) controls cytoplasmic events during virion maturation
- PMID: 16873276
- PMCID: PMC1563810
- DOI: 10.1128/JVI.00457-06
Betaherpesvirus-conserved cytomegalovirus tegument protein ppUL32 (pp150) controls cytoplasmic events during virion maturation
Abstract
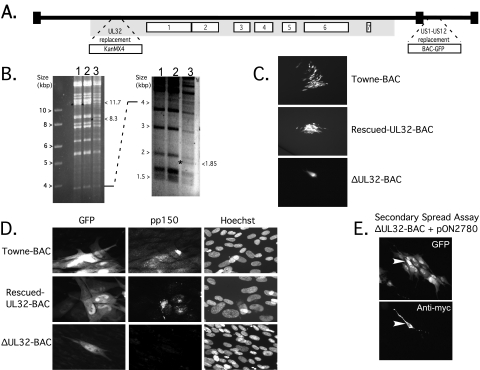
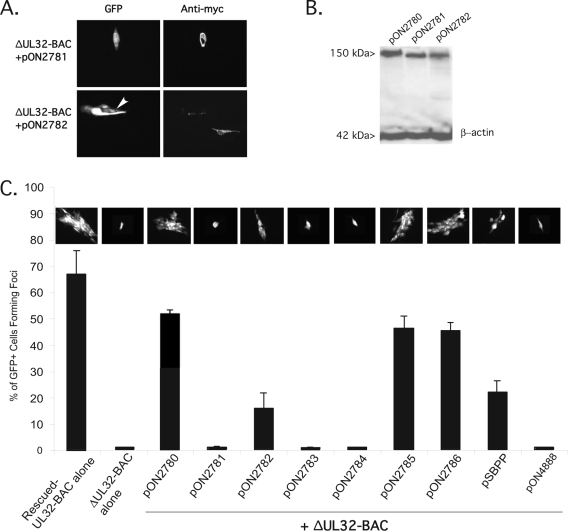
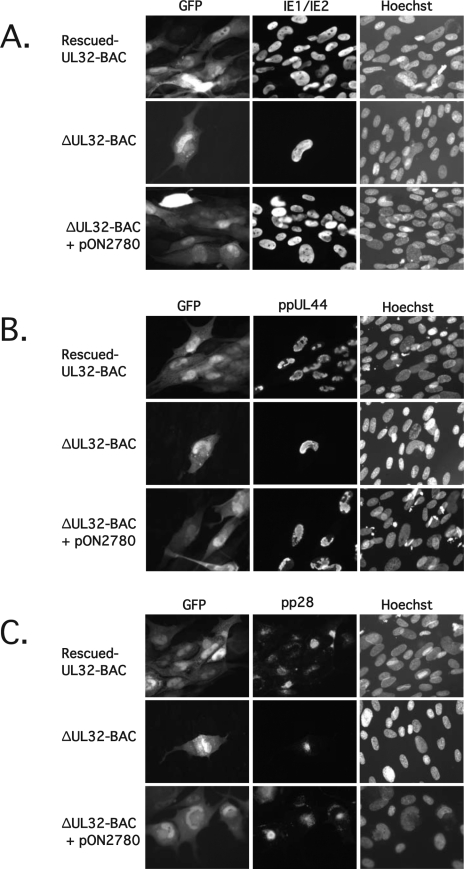
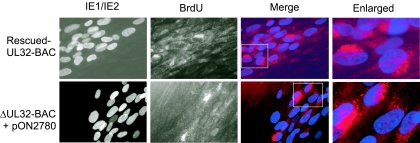
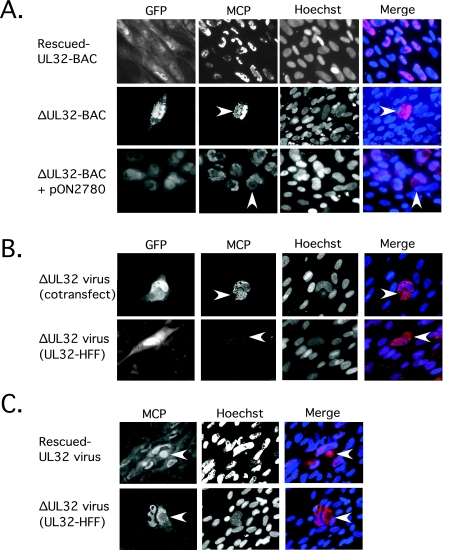
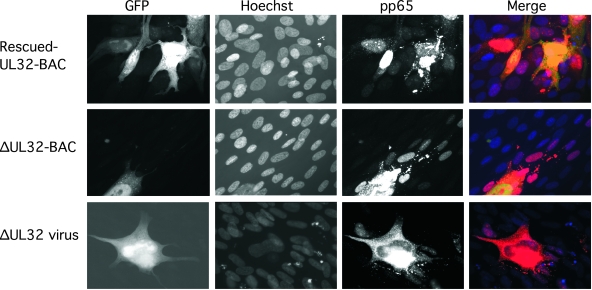
The UL32 gene of human cytomegalovirus (CMV) encodes a prominent betaherpesvirus-conserved virion tegument protein, called pp150 (basic phosphoprotein/ppUL32), that accumulates within a cytoplasmic inclusion adjacent to the nucleus at late times during infection. Using a UL32 deletion mutant (DeltaUL32-BAC) (where BAC is bacterial artificial chromosome), we demonstrate that pp150 is critical for virion maturation in the cytoplasmic compartment. Cotransfection of a pp150 expression plasmid with DeltaUL32-BAC DNA led to complementation of the replication defect with focus formation due to secondary spread. Deletion of the amino terminus of pp150 or disruption of the betaherpesvirus conserved regions, CR1 and CR2, revealed these regions to be critical for replication. In contrast, deletion of the carboxyl terminus only partially compromised maturation while disruption of glycosylation sites had no effect. An African green monkey CMV UL32 homolog complemented DeltaUL32-BAC replication but murine CMV M32 failed to complement, consistent with evolutionary divergence of rodent and primate cytomegaloviruses. Infection with DeltaUL32-BAC showed normal expression of all kinetic classes of viral genes and replication of viral DNA, with accumulation of viral DNA-containing particles in the cytoplasm; however, mutant virus did not spread to adjacent cells. In contrast to this block in virion infectivity, cell-to-cell transfer of pp65-containing particles was observed, suggesting that release of dense bodies continued in the absence of pp150. These observations demonstrate that pp150 is critical for virion egress, possibly at the stage of final envelopment.
Figures

Similar articles
-
Control of cytoplasmic maturation events by cytomegalovirus tegument protein pp150.J Virol. 2008 Oct;82(19):9433-44. doi: 10.1128/JVI.00533-08. Epub 2008 Jul 23. J Virol. 2008. PMID: 18653449 Free PMC article.
-
Accumulation of virion tegument and envelope proteins in a stable cytoplasmic compartment during human cytomegalovirus replication: characterization of a potential site of virus assembly.J Virol. 2000 Jan;74(2):975-86. doi: 10.1128/jvi.74.2.975-986.2000. J Virol. 2000. PMID: 10623760 Free PMC article.
-
Nuclear localization of the human cytomegalovirus tegument protein pp150 (ppUL32).J Gen Virol. 1995 Jul;76 ( Pt 7):1591-601. doi: 10.1099/0022-1317-76-7-1591. J Gen Virol. 1995. PMID: 9049366
-
[Structure and assembly of human beta herpesviruses].Nihon Rinsho. 1998 Jan;56(1):22-8. Nihon Rinsho. 1998. PMID: 9465660 Review. Japanese.
-
Betaherpesvirus assembly and egress: Recent advances illuminate the path.Adv Virus Res. 2020;108:337-392. doi: 10.1016/bs.aivir.2020.09.003. Epub 2020 Oct 7. Adv Virus Res. 2020. PMID: 33837722 Review.
Cited by
-
The A, B, Cs of herpesvirus capsids.Viruses. 2015 Feb 26;7(3):899-914. doi: 10.3390/v7030899. Viruses. 2015. PMID: 25730559 Free PMC article. Review.
-
Human Cytomegalovirus Primary Infection and Reactivation: Insights From Virion-Carried Molecules.Front Microbiol. 2020 Jul 14;11:1511. doi: 10.3389/fmicb.2020.01511. eCollection 2020. Front Microbiol. 2020. PMID: 32765441 Free PMC article. Review.
-
Human Cytomegalovirus pUL47 Modulates Tegumentation and Capsid Accumulation at the Viral Assembly Complex.J Virol. 2015 Jul;89(14):7314-28. doi: 10.1128/JVI.00603-15. Epub 2015 May 6. J Virol. 2015. PMID: 25948747 Free PMC article.
-
Bioinformatics and Functional Analysis of a New Nuclear Localization Sequence of the Influenza A Virus Nucleoprotein.Cells. 2022 Sep 22;11(19):2957. doi: 10.3390/cells11192957. Cells. 2022. PMID: 36230922 Free PMC article.
-
Viral and host control of cytomegalovirus maturation.Trends Microbiol. 2012 Aug;20(8):392-401. doi: 10.1016/j.tim.2012.04.008. Epub 2012 May 23. Trends Microbiol. 2012. PMID: 22633075 Free PMC article. Review.
References
-
- Bartz, S. R., and M. A. Vodicka. 1997. Production of high-titer human immunodeficiency virus type 1 pseudotyped with vesicular stomatitis virus glycoprotein. Methods 12:337-342. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
