

An early role for sonic hedgehog from foregut endoderm in jaw development: ensuring neural crest cell survival
- PMID: 16868080
- PMCID: PMC1544217
- DOI: 10.1073/pnas.0604751103
An early role for sonic hedgehog from foregut endoderm in jaw development: ensuring neural crest cell survival
Abstract
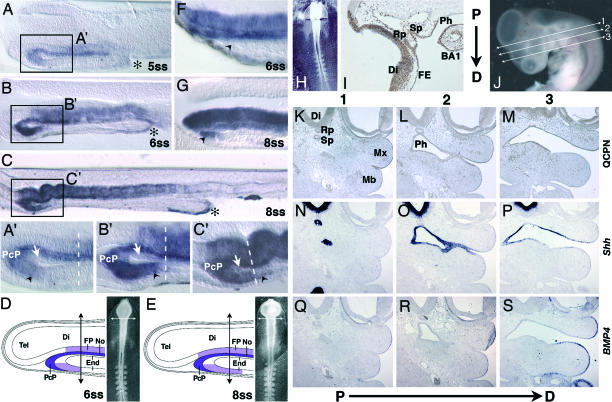
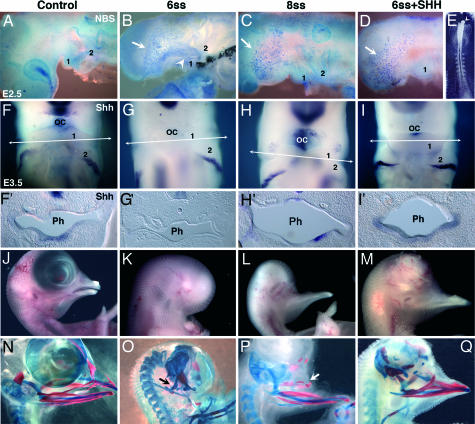
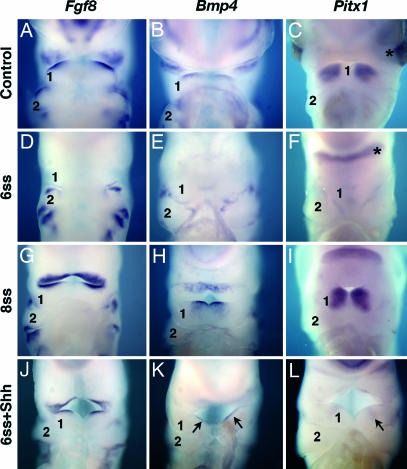
We have investigated the role of Sonic hedgehog (Shh) in the development of facial structures by depriving chicken embryos of the most anterior sources of this morphogen, including the prechordal plate and the anterior ventral endoderm of the foregut, before the onset of neural crest cell (NCC) migration to the first branchial arch (BA1). The entire forehead, including the foregut endoderm, was removed at 5- to 10-somite stage (ss), which led to the absence of the lower jaw when the operation was performed before 7-ss. If the embryos were deprived of their forehead at 8- to 10-ss, they were later on endowed with a lower beak. In embryos that were operated on early, the NCCs migrated normally to BA1 but were subjected to massive apoptosis a few hours later. Cell death did not occur when forehead excision was performed at a later stage. In this case, onward expression of Shh in the ventral foregut endoderm extended caudally over the excision limit, and we hypothesized that absence of Shh production by the endoderm in embryos that were operated on early could be responsible for the NCC apoptosis and the failure of BA1 development. We thus provided exogenous Shh to the embryos that were operated on before 7-ss. In this case, the development of the lower jaw was rescued. Therefore, Shh derived from the ventral foregut endoderm ensures the survival of NCCs at a critical stage of BA1 development.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
Similar articles
-
Sonic Hedgehog promotes the survival of neural crest cells by limiting apoptosis induced by the dependence receptor CDON during branchial arch development.Biochem Biophys Res Commun. 2014 Sep 26;452(3):655-60. doi: 10.1016/j.bbrc.2014.08.134. Epub 2014 Sep 1. Biochem Biophys Res Commun. 2014. PMID: 25193697
-
Induction of mirror-image supernumerary jaws in chicken mandibular mesenchyme by Sonic Hedgehog-producing cells.Development. 2008 Jul;135(13):2311-9. doi: 10.1242/dev.019125. Development. 2008. PMID: 18539924
-
Patterning of the hyoid cartilage depends upon signals arising from the ventral foregut endoderm.Dev Dyn. 2003 Oct;228(2):239-46. doi: 10.1002/dvdy.10380. Dev Dyn. 2003. PMID: 14517995
-
Craniofacial development: discoveries made in the chicken embryo.Int J Dev Biol. 2018;62(1-2-3):97-107. doi: 10.1387/ijdb.170321ja. Int J Dev Biol. 2018. PMID: 29616744 Review.
-
About face: signals and genes controlling jaw patterning and identity in vertebrates.Bioessays. 2003 Jun;25(6):554-68. doi: 10.1002/bies.10288. Bioessays. 2003. PMID: 12766945 Review.
Cited by
-
Diversity in primary palate ontogeny of amniotes revealed with 3D imaging.J Anat. 2015 May;226(5):420-33. doi: 10.1111/joa.12291. Epub 2015 Apr 22. J Anat. 2015. PMID: 25904546 Free PMC article.
-
Endoderm-derived Sonic hedgehog and mesoderm Hand2 expression are required for enteric nervous system development in zebrafish.Dev Biol. 2008 Jun 1;318(1):52-64. doi: 10.1016/j.ydbio.2008.02.061. Epub 2008 Mar 20. Dev Biol. 2008. PMID: 18436202 Free PMC article.
-
High frequency of cephalic neural crest cells shows coexistence of neurogenic, melanogenic, and osteogenic differentiation capacities.Proc Natl Acad Sci U S A. 2009 Jun 2;106(22):8947-52. doi: 10.1073/pnas.0903780106. Epub 2009 May 15. Proc Natl Acad Sci U S A. 2009. PMID: 19447928 Free PMC article.
-
Congenital aural atresia associated with agenesis of internal carotid artery in a girl with a FOXI3 deletion.Am J Med Genet A. 2015 Mar;167A(3):537-44. doi: 10.1002/ajmg.a.36895. Epub 2015 Feb 5. Am J Med Genet A. 2015. PMID: 25655429 Free PMC article.
-
Thyroid C-Cell Biology and Oncogenic Transformation.Recent Results Cancer Res. 2015;204:1-39. doi: 10.1007/978-3-319-22542-5_1. Recent Results Cancer Res. 2015. PMID: 26494382 Free PMC article. Review.
References
-
- Couly G., Grapin-Botton A., Coltey P., Le Douarin N. M. Development (Cambridge, U.K.) 1996;122:3393–3407. - PubMed
-
- Kontges G., Lumsden A. Development (Cambridge, U.K.) 1996;122:3229–3242. - PubMed
-
- Le Douarin N. M., Kalcheim C. The Neural Crest. Cambridge, U.K.: Cambridge Univ. Press; 1999.
-
- Etchevers H. C., Couly G., Vincent C., Le Douarin N. M. Development (Cambridge, U.K.) 1999;126:3533–3543. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
