

Interaction of survival and death signaling in basal forebrain neurons: roles of neurotrophins and proneurotrophins
- PMID: 16855103
- PMCID: PMC6674285
- DOI: 10.1523/JNEUROSCI.1560-06.2006
Interaction of survival and death signaling in basal forebrain neurons: roles of neurotrophins and proneurotrophins
Abstract
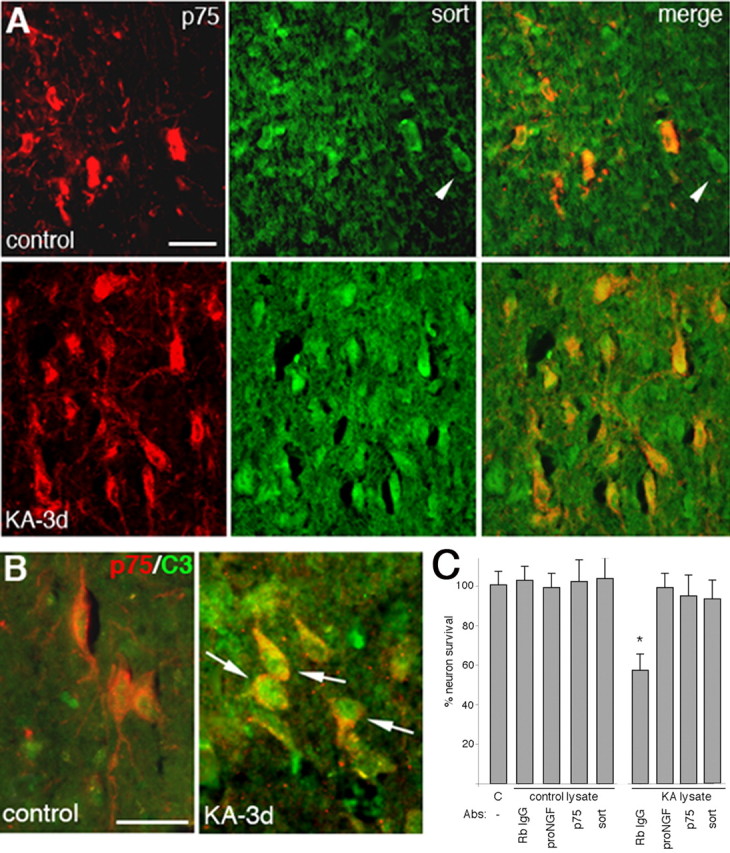
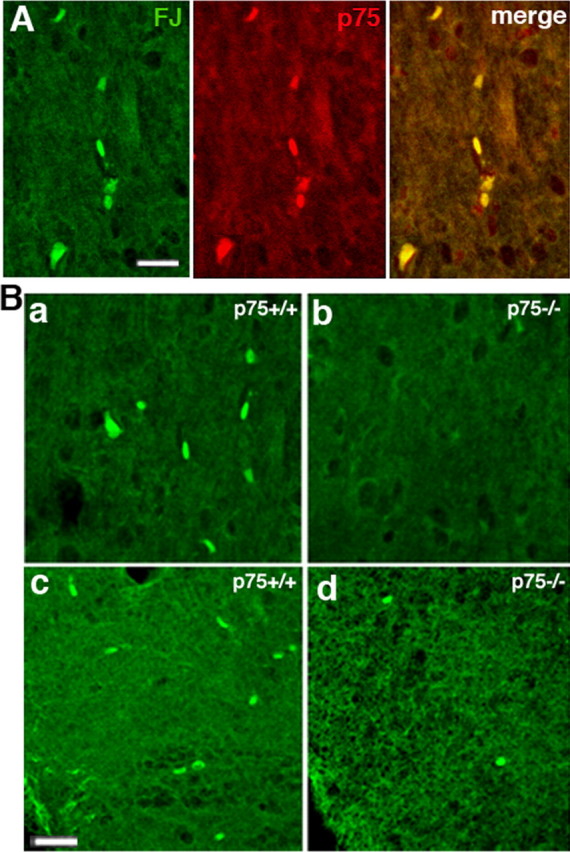
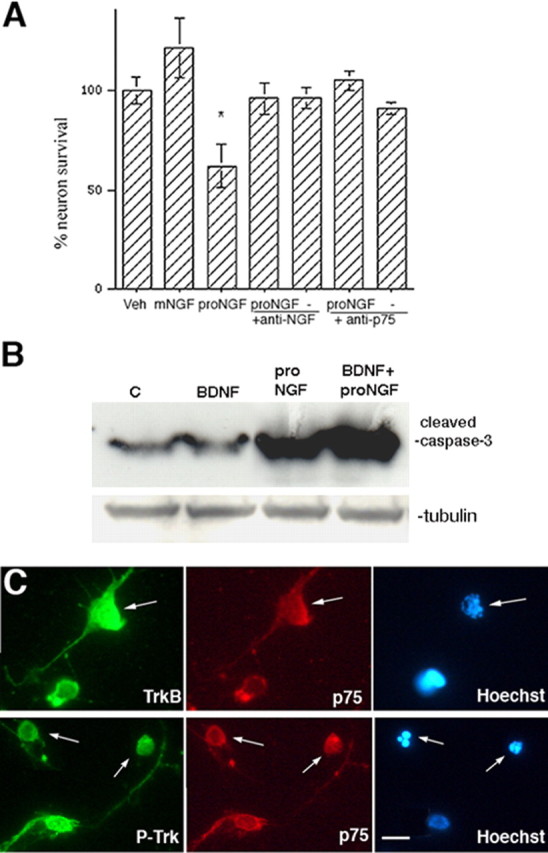
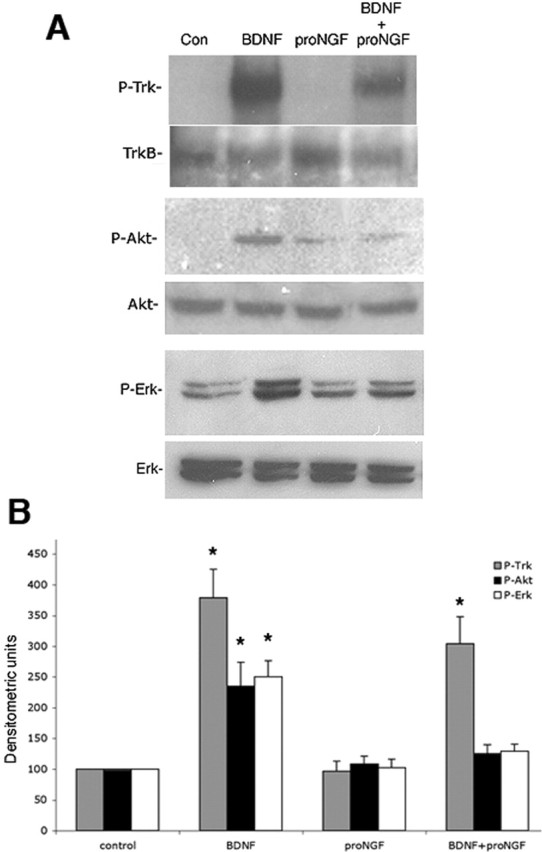
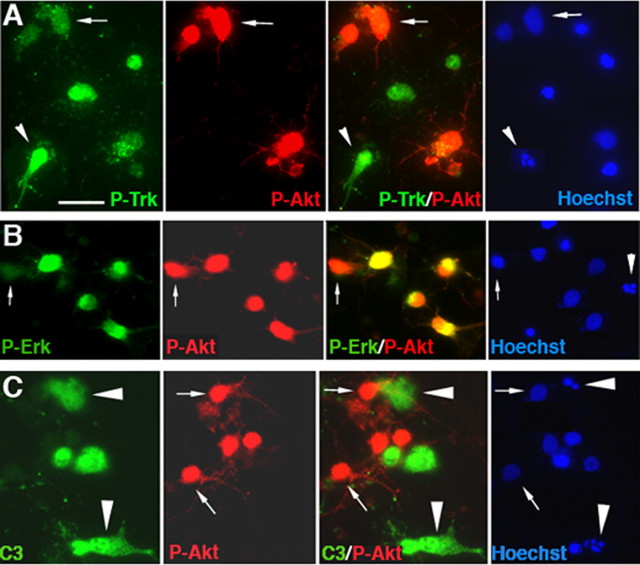
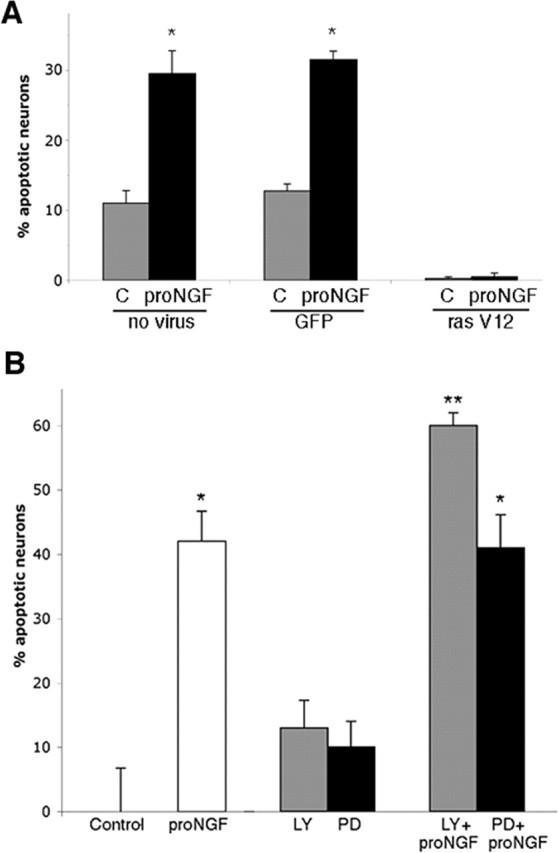
Proneurotrophins bind with high affinity to p75 neurotrophin receptor (p75NTR) and lack the capacity to bind Trk receptors, suggesting that proneurotrophins can elicit apoptosis via p75NTR even in cells expressing survival-promoting Trk receptors. In the CNS, basal forebrain (BF) neurons are particularly vulnerable to degeneration in Alzheimer's disease, and are among the few populations of brain neurons that express p75NTR throughout life. These neurons also express Trk receptors and may be concomitantly exposed to both proneurotrophins and mature neurotrophins during development, disease, or after injury. We investigated the interaction of mature and proneurotrophin signaling in these CNS neurons. Kainic acid-induced seizures elicited production of pro-NGF by BF astrocytes before caspase activation in p75NTR-positive BF neurons, demonstrating local production of proneurotrophins under pathological conditions and suggesting apoptotic signaling in vivo. Mechanisms of proneurotrophin-induced death were analyzed in cultured BF neurons, and required both p75NTR and its coreceptor sortilin. Surprisingly, exposure to both mature neurotrophins and proneurotrophins demonstrated that Trk phosphorylation did not prevent pro-NGF-induced apoptosis via p75NTR. However, activation of PI3K (phosphatidylinositol 3-kinase)/Akt and MEK (mitogen-activated protein kinase kinase)/Erk pathways prevented pro-NGF-induced apoptosis, revealing a novel critical checkpoint in survival versus apoptotic signaling downstream of Trk activation, and suggesting that pro-NGF blocks survival signaling by preventing Akt and Erk activation. This study shows that proneurotrophins are produced in the brain under pathological conditions, and can elicit apoptosis of BF neurons even when Trk receptors are activated.
Figures




Similar articles
-
Pro-NGF, sortilin, and p75NTR: potential mediators of injury-induced apoptosis in the mouse dorsal root ganglion.Brain Res. 2007 Dec 5;1183:32-42. doi: 10.1016/j.brainres.2007.09.051. Epub 2007 Oct 26. Brain Res. 2007. PMID: 17964555 Free PMC article.
-
ProNGF induces PTEN via p75NTR to suppress Trk-mediated survival signaling in brain neurons.J Neurosci. 2010 Nov 17;30(46):15608-15. doi: 10.1523/JNEUROSCI.2581-10.2010. J Neurosci. 2010. PMID: 21084616 Free PMC article.
-
ProBDNF induces neuronal apoptosis via activation of a receptor complex of p75NTR and sortilin.J Neurosci. 2005 Jun 1;25(22):5455-63. doi: 10.1523/JNEUROSCI.5123-04.2005. J Neurosci. 2005. PMID: 15930396 Free PMC article.
-
Proneurotrophins, seizures, and neuronal apoptosis.Neuroscientist. 2010 Jun;16(3):244-52. doi: 10.1177/1073858409349903. Epub 2010 Apr 1. Neuroscientist. 2010. PMID: 20360602 Free PMC article. Review.
-
Understanding proneurotrophin actions: Recent advances and challenges.Dev Neurobiol. 2010 Apr;70(5):350-9. doi: 10.1002/dneu.20768. Dev Neurobiol. 2010. PMID: 20186707 Free PMC article. Review.
Cited by
-
Impact of the NGF maturation and degradation pathway on the cortical cholinergic system phenotype.J Neurosci. 2012 Feb 8;32(6):2002-12. doi: 10.1523/JNEUROSCI.1144-11.2012. J Neurosci. 2012. PMID: 22323714 Free PMC article.
-
RhoGDI phosphorylation by PKC promotes its interaction with death receptor p75NTR to gate axon growth and neuron survival.EMBO Rep. 2024 Mar;25(3):1490-1512. doi: 10.1038/s44319-024-00064-2. Epub 2024 Jan 22. EMBO Rep. 2024. PMID: 38253689 Free PMC article.
-
Pro-NGF, sortilin, and p75NTR: potential mediators of injury-induced apoptosis in the mouse dorsal root ganglion.Brain Res. 2007 Dec 5;1183:32-42. doi: 10.1016/j.brainres.2007.09.051. Epub 2007 Oct 26. Brain Res. 2007. PMID: 17964555 Free PMC article.
-
Neurotrophin receptors expression and JNK pathway activation in human astrocytomas.BMC Cancer. 2007 Oct 31;7:202. doi: 10.1186/1471-2407-7-202. BMC Cancer. 2007. PMID: 17971243 Free PMC article.
-
Pathophysiological mechanisms for actions of the neurotrophins.Brain Pathol. 2006 Oct;16(4):320-32. doi: 10.1111/j.1750-3639.2006.00039.x. Brain Pathol. 2006. PMID: 17107602 Free PMC article. Review.
References
-
- Alderson RF, Alterman AL, Barde Y-A, Lindsay RM (1990). Brain-derived neurotrophic factor increases survival and differentiated functions of rat septal cholinergic neurons in culture. Neuron 5:297–306. - PubMed
-
- Barker PA (2004). p75NTR is positively promiscuous: novel partners and new insights. Neuron 42:529–533. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous