

"Self" and "nonself" manipulation of interferon defense during persistent infection: bovine viral diarrhea virus resists alpha/beta interferon without blocking antiviral activity against unrelated viruses replicating in its host cells
- PMID: 16809298
- PMCID: PMC1489018
- DOI: 10.1128/JVI.02443-05
"Self" and "nonself" manipulation of interferon defense during persistent infection: bovine viral diarrhea virus resists alpha/beta interferon without blocking antiviral activity against unrelated viruses replicating in its host cells
Abstract
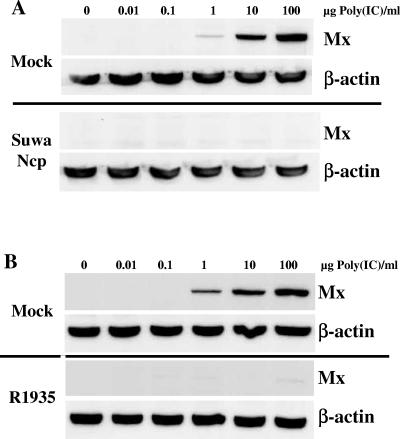
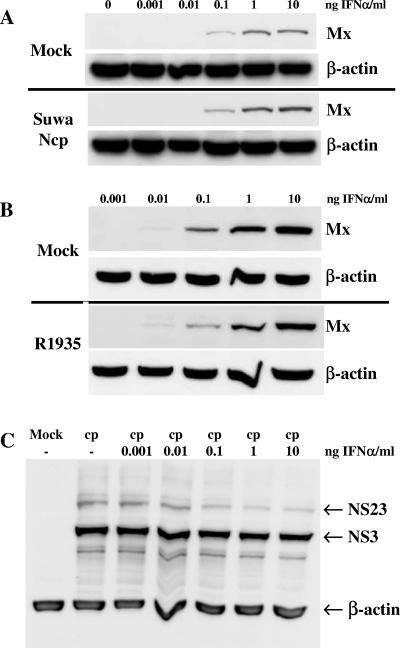
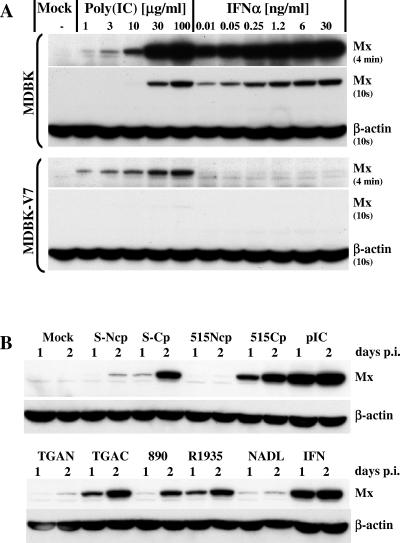
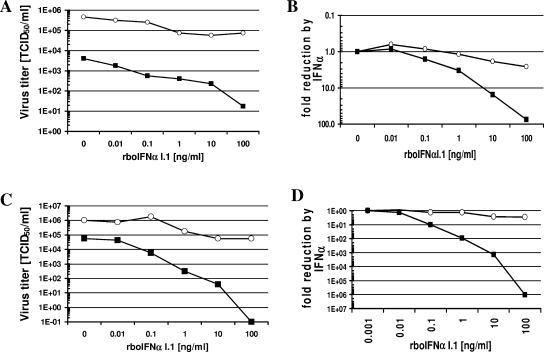
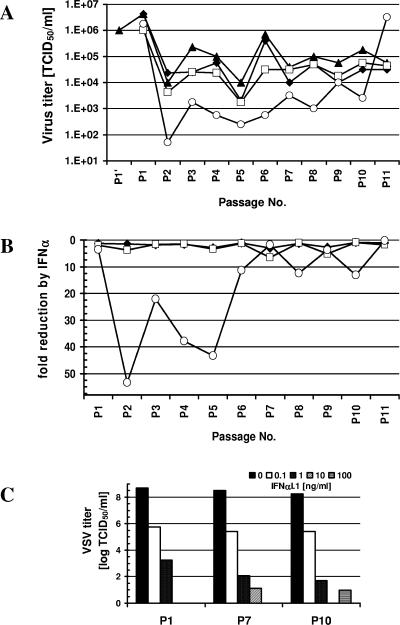
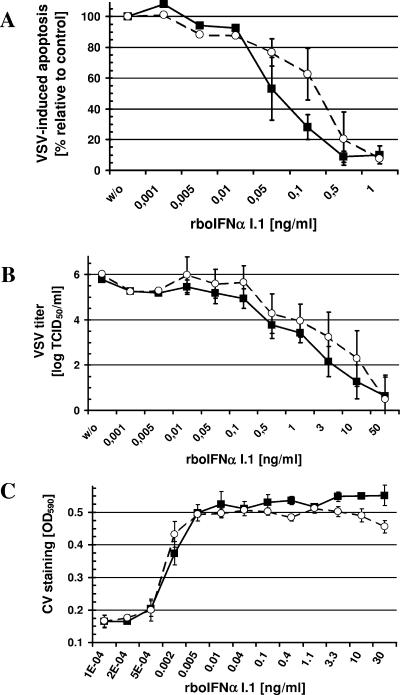
Bovine viral diarrhea virus (BVDV), together with Classical swine fever virus (CSFV) and Border disease virus (BDV) of sheep, belongs to the genus Pestivirus of the Flaviviridae. BVDV is either cytopathic (cp) or noncytopathic (ncp), as defined by its effect on cultured cells. Infection of pregnant animals with the ncp biotype may lead to the birth of persistently infected calves that are immunotolerant to the infecting viral strain. In addition to evading the adaptive immune system, BVDV evades key mechanisms of innate immunity. Previously, we showed that ncp BVDV inhibits the induction of apoptosis and alpha/beta interferon (IFN-alpha/beta) synthesis by double-stranded RNA (dsRNA). Here, we report that (i) both ncp and cp BVDV block the induction by dsRNA of the Mx protein (which can also be induced in the absence of IFN signaling); (ii) neither biotype blocks the activity of IFN; and (iii) once infection is established, BVDV is largely resistant to the activity of IFN-alpha/beta but (iv) does not interfere with the establishment of an antiviral state induced by IFN-alpha/beta against unrelated viruses. The results of our study suggest that, in persistent infection, BVDV is able to evade a central element of innate immunity directed against itself without generally compromising its activity against unrelated viruses ("nonself") that may replicate in cells infected with ncp BVDV. This highly selective "self" and "nonself" model of evasion of the interferon defense system may be a key element in the success of persistent infection in addition to immunotolerance initiated by the early time point of fetal infection.
Figures
Similar articles
-
Noncytopathic bovine viral diarrhea virus inhibits double-stranded RNA-induced apoptosis and interferon synthesis.J Virol. 2001 May;75(10):4692-8. doi: 10.1128/JVI.75.10.4692-4698.2001. J Virol. 2001. PMID: 11312340 Free PMC article.
-
Induction of interferon-gamma and downstream pathways during establishment of fetal persistent infection with bovine viral diarrhea virus.Virus Res. 2014 Apr;183:95-106. doi: 10.1016/j.virusres.2014.02.002. Epub 2014 Feb 12. Virus Res. 2014. PMID: 24530541
-
Inhibition of beta interferon transcription by noncytopathogenic bovine viral diarrhea virus is through an interferon regulatory factor 3-dependent mechanism.J Virol. 2002 Sep;76(18):8979-88. doi: 10.1128/jvi.76.18.8979-8988.2002. J Virol. 2002. PMID: 12186882 Free PMC article.
-
[Bovine diarrhea virus: an update].Rev Argent Microbiol. 1997 Jan-Mar;29(1):47-61. Rev Argent Microbiol. 1997. PMID: 9229725 Review. Spanish.
-
[How the bovine viral diarrhea virus outwits the immune system].Dtsch Tierarztl Wochenschr. 2006 Apr;113(4):124-9. Dtsch Tierarztl Wochenschr. 2006. PMID: 16716045 Review. German.
Cited by
-
Genetic characterization of bovine viral diarrhea virus strains in Beijing, China and innate immune responses of peripheral blood mononuclear cells in persistently infected dairy cattle.J Vet Sci. 2015;16(4):491-500. doi: 10.4142/jvs.2015.16.4.491. J Vet Sci. 2015. PMID: 26119170 Free PMC article.
-
Mitochondria-mediated ferroptosis contributes to the inflammatory responses of bovine viral diarrhea virus (BVDV) in vitro.J Virol. 2024 Feb 20;98(2):e0188023. doi: 10.1128/jvi.01880-23. Epub 2024 Jan 16. J Virol. 2024. PMID: 38226812 Free PMC article.
-
Epigenomic and Proteomic Changes in Fetal Spleens Persistently Infected with Bovine Viral Diarrhea Virus: Repercussions for the Developing Immune System, Bone, Brain, and Heart.Viruses. 2022 Feb 28;14(3):506. doi: 10.3390/v14030506. Viruses. 2022. PMID: 35336913 Free PMC article.
-
Transmission of bovine viral diarrhea virus among white-tailed deer (Odocoileus virginianus).Vet Res. 2010 Mar-Apr;41(2):20. doi: 10.1051/vetres/2009068. Epub 2009 Nov 19. Vet Res. 2010. PMID: 19922743 Free PMC article.
-
Persistent bovine viral diarrhea virus (BVDV) infection in cattle herds.Iran J Vet Res. 2017 Summer;18(3):154-163. Iran J Vet Res. 2017. PMID: 29163643 Free PMC article. Review.
References
-
- Adler, B., H. Adler, T. W. Jungi, and E. Peterhans. 1995. Interferon-alpha primes macrophages for lipopolysaccharide-induced apoptosis. Biochem. Biophys. Res. Commun. 215:921-927. - PubMed
-
- Adler, H., B. Frech, P. Meier, T. W. Jungi, and E. Peterhans. 1994. Noncytopathic strains of bovine viral diarrhea virus prime bovine bone marrow-derived macrophages for enhanced generation of nitric oxide. Biochem. Biophys. Res. Commun. 202:1562-1568. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
