

Structure and E3-ligase activity of the Ring-Ring complex of polycomb proteins Bmi1 and Ring1b
- PMID: 16710298
- PMCID: PMC1478191
- DOI: 10.1038/sj.emboj.7601144
Structure and E3-ligase activity of the Ring-Ring complex of polycomb proteins Bmi1 and Ring1b
Abstract
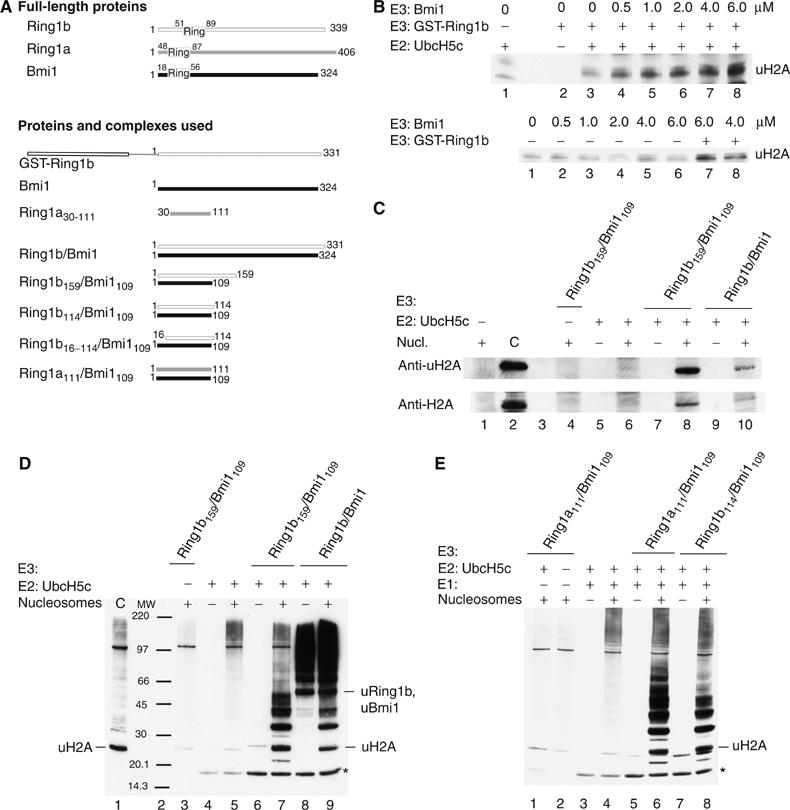
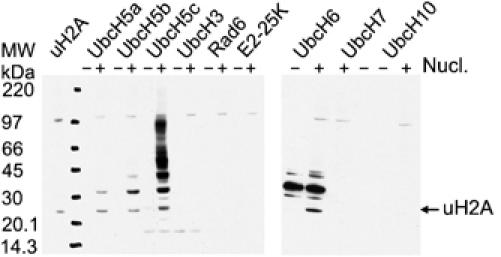
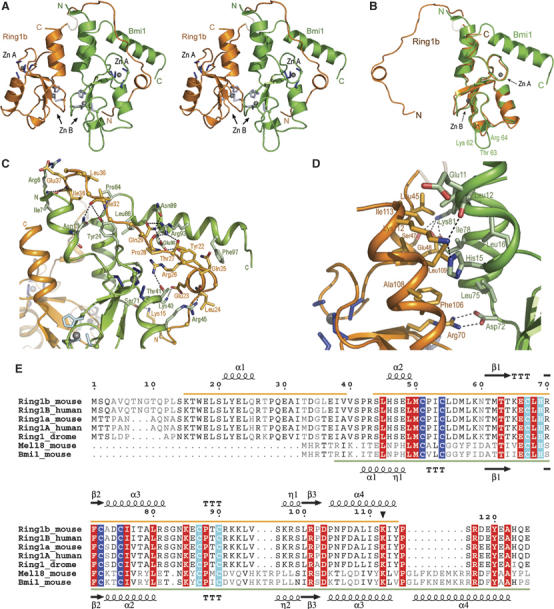
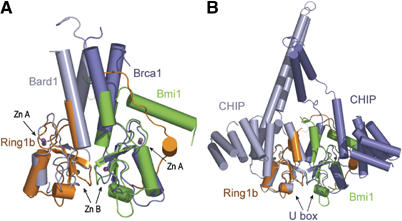
Polycomb group proteins Ring1b and Bmi1 (B-cell-specific Moloney murine leukaemia virus integration site 1) are critical components of the chromatin modulating PRC1 complex. Histone H2A ubiquitination by the PRC1 complex strongly depends on the Ring1b protein. Here we show that the E3-ligase activity of Ring1b on histone H2A is enhanced by Bmi1 in vitro. The N-terminal Ring-domains are sufficient for this activity and Ring1a can replace Ring1b. E2 enzymes UbcH5a, b, c or UbcH6 support this activity with varying processivity and selectivity. All four E2s promote autoubiquitination of Ring1b without affecting E3-ligase activity. We solved the crystal structure of the Ring-Ring heterodimeric complex of Ring1b and Bmi1. In the structure the arrangement of the Ring-domains is similar to another H2A E3 ligase, the BRCA1/BARD1 complex, but complex formation depends on an N-terminal arm of Ring1b that embraces the Bmi1 Ring-domain. Mutation of a critical residue in the E2/E3 interface shows that catalytic activity resides in Ring1b and not in Bmi1. These data provide a foundation for understanding the critical enzymatic activity at the core of the PRC1 polycomb complex, which is implicated in stem cell maintenance and cancer.
Figures


Similar articles
-
Recognition of UbcH5c and the nucleosome by the Bmi1/Ring1b ubiquitin ligase complex.EMBO J. 2011 Jul 19;30(16):3285-97. doi: 10.1038/emboj.2011.243. EMBO J. 2011. PMID: 21772249 Free PMC article.
-
The polycomb protein Ring1B generates self atypical mixed ubiquitin chains required for its in vitro histone H2A ligase activity.Mol Cell. 2006 Dec 8;24(5):701-711. doi: 10.1016/j.molcel.2006.10.022. Mol Cell. 2006. PMID: 17157253
-
Structure of a Bmi-1-Ring1B polycomb group ubiquitin ligase complex.J Biol Chem. 2006 Jul 21;281(29):20643-9. doi: 10.1074/jbc.M602461200. Epub 2006 May 18. J Biol Chem. 2006. PMID: 16714294
-
Writing Histone Monoubiquitination in Human Malignancy-The Role of RING Finger E3 Ubiquitin Ligases.Genes (Basel). 2019 Jan 18;10(1):67. doi: 10.3390/genes10010067. Genes (Basel). 2019. PMID: 30669413 Free PMC article. Review.
-
Role of polycomb proteins Ring1A and Ring1B in the epigenetic regulation of gene expression.Int J Dev Biol. 2009;53(2-3):355-70. doi: 10.1387/ijdb.082690mv. Int J Dev Biol. 2009. PMID: 19412891 Review.
Cited by
-
Critical role of cysteine-266 of SIE3 in regulating the ubiquitination and degradation of SIP1 transcription factor in Lotus japonicus.Planta. 2021 May 25;253(6):126. doi: 10.1007/s00425-021-03647-8. Planta. 2021. PMID: 34036431
-
Estrogen induces dynamic ERα and RING1B recruitment to control gene and enhancer activities in luminal breast cancer.Sci Adv. 2020 Jun 5;6(23):eaaz7249. doi: 10.1126/sciadv.aaz7249. eCollection 2020 Jun. Sci Adv. 2020. PMID: 32548262 Free PMC article.
-
Crosstalk among Epigenetic Pathways Regulates Neurogenesis.Front Neurosci. 2012 May 8;6:59. doi: 10.3389/fnins.2012.00059. eCollection 2012. Front Neurosci. 2012. PMID: 22586361 Free PMC article.
-
The Hemogenic Competence of Endothelial Progenitors Is Restricted by Runx1 Silencing during Embryonic Development.Cell Rep. 2016 Jun 7;15(10):2185-2199. doi: 10.1016/j.celrep.2016.05.001. Epub 2016 May 26. Cell Rep. 2016. PMID: 27239041 Free PMC article.
-
HECT and RING finger families of E3 ubiquitin ligases at a glance.J Cell Sci. 2012 Feb 1;125(Pt 3):531-7. doi: 10.1242/jcs.091777. J Cell Sci. 2012. PMID: 22389392 Free PMC article. Review. No abstract available.
References
-
- Abrahams JP, Leslie AG (1996) Methods used in the structure determination of bovine mitochondrial F1 ATPase. Acta Crystallogr D 52: 30–42 - PubMed
-
- Alkema MJ, Jacobs H, van Lohuizen M, Berns A (1997) Pertubation of B and T cell development and predisposition to lymphomagenesis in Emu Bmi1 transgenic mice require the Bmi1 RING finger. Oncogene 15: 899–910 - PubMed
-
- Andersen P, Kragelund BB, Olsen AN, Larsen FH, Chua NH, Poulsen FM, Skriver K (2004) Structure and biochemical function of a prototypical Arabidopsis U-box domain. J Biol Chem 279: 40053–40061 - PubMed
-
- Bellon SF, Rodgers KK, Schatz DG, Coleman JE, Steitz TA (1997) Crystal structure of the RAG1 dimerization domain reveals multiple zinc-binding motifs including a novel zinc binuclear cluster. Nat Struct Biol 4: 586–591 - PubMed
-
- Borden KL (2000) Ring-domains: master builders of molecular scaffolds? J Mol Biol 295: 1103–1112 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
