

Systematic identification and functional screens of uncharacterized proteins associated with eukaryotic ribosomal complexes
- PMID: 16702403
- PMCID: PMC1472904
- DOI: 10.1101/gad.1422006
Systematic identification and functional screens of uncharacterized proteins associated with eukaryotic ribosomal complexes
Abstract
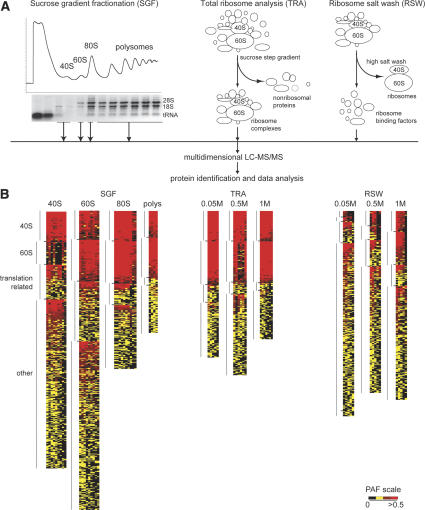
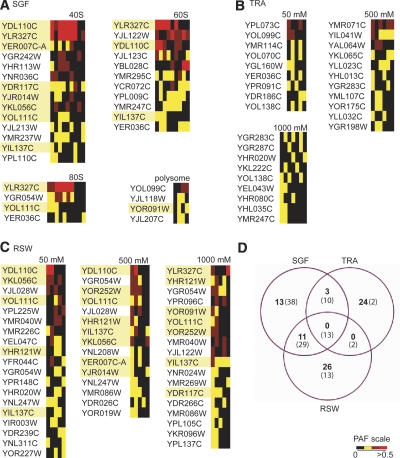
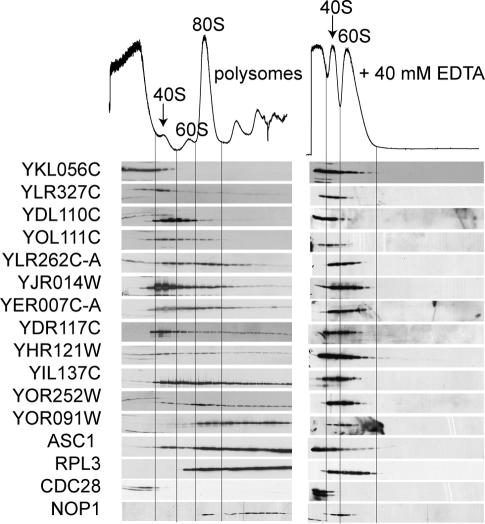
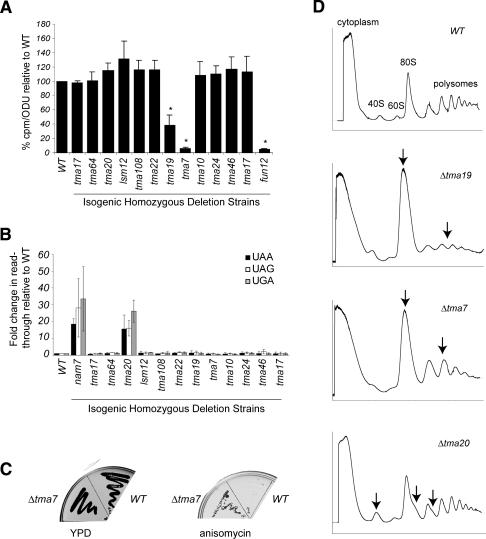
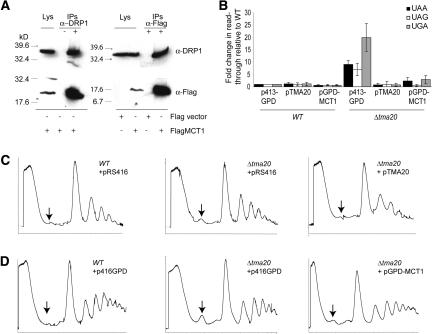
Translation regulation is a critical means by which cells control growth, division, and apoptosis. To gain further insight into translation and related processes, we performed multifaceted mass spectrometry-based proteomic screens of yeast ribosomal complexes and discovered an association of 77 uncharacterized yeast proteins with ribosomes. Immunoblotting revealed an EDTA-dependent cosedimentation with ribosomes in sucrose gradients for 11 candidate translation-machinery-associated (TMA) proteins. Tandem affinity purification linked one candidate, LSM12, to the RNA processing proteins PBP1 and PBP4. A second candidate, TMA46, interacted with RBG1, a GTPase that interacts with ribosomes. By adapting translation assays to high-throughput screening methods, we showed that null yeast strains harboring deletions for several of the TMA genes had alterations in protein synthesis rates (TMA7 and TMA19), susceptibility to drugs that inhibit translation (TMA7), translation fidelity (TMA20), and polyribosome profiles (TMA7, TMA19, and TMA20). TMA20 has significant sequence homology with the oncogene MCT-1. Expression of human MCT-1 in the Deltatma20 yeast mutant complemented translation-related defects, strongly implying that MCT-1 functions in translation-related processes. Together these findings implicate the TMA proteins and, potentially, their human homologs, in translation related processes.
Figures
Similar articles
-
40S ribosome profiling reveals distinct roles for Tma20/Tma22 (MCT-1/DENR) and Tma64 (eIF2D) in 40S subunit recycling.Nat Commun. 2021 May 20;12(1):2976. doi: 10.1038/s41467-021-23223-8. Nat Commun. 2021. PMID: 34016977 Free PMC article.
-
Multi-protein bridging factor 1(Mbf1), Rps3 and Asc1 prevent stalled ribosomes from frameshifting.Elife. 2018 Nov 22;7:e39637. doi: 10.7554/eLife.39637. Elife. 2018. PMID: 30465652 Free PMC article.
-
Ribosomal protein L33 is required for ribosome biogenesis, subunit joining, and repression of GCN4 translation.Mol Cell Biol. 2007 Sep;27(17):5968-85. doi: 10.1128/MCB.00019-07. Epub 2007 Jun 4. Mol Cell Biol. 2007. PMID: 17548477 Free PMC article.
-
Ribosome Stoichiometry: From Form to Function.Trends Biochem Sci. 2019 Feb;44(2):95-109. doi: 10.1016/j.tibs.2018.10.009. Epub 2018 Nov 22. Trends Biochem Sci. 2019. PMID: 30473427 Free PMC article. Review.
-
Ribosome synthesis meets the cell cycle.Curr Opin Microbiol. 2004 Dec;7(6):631-7. doi: 10.1016/j.mib.2004.10.007. Curr Opin Microbiol. 2004. PMID: 15556036 Review.
Cited by
-
Evolving precision: rRNA expansion segment 7S modulates translation velocity and accuracy in eukaryal ribosomes.Nucleic Acids Res. 2024 Apr 24;52(7):4021-4036. doi: 10.1093/nar/gkae067. Nucleic Acids Res. 2024. PMID: 38324474 Free PMC article.
-
Pbp1 is involved in Ccr4- and Khd1-mediated regulation of cell growth through association with ribosomal proteins Rpl12a and Rpl12b.Eukaryot Cell. 2013 Jun;12(6):864-74. doi: 10.1128/EC.00370-12. Epub 2013 Apr 5. Eukaryot Cell. 2013. PMID: 23563484 Free PMC article.
-
Localization to, and effects of Pbp1, Pbp4, Lsm12, Dhh1, and Pab1 on stress granules in Saccharomyces cerevisiae.PLoS One. 2010 Apr 2;5(4):e10006. doi: 10.1371/journal.pone.0010006. PLoS One. 2010. PMID: 20368989 Free PMC article.
-
A genome-wide screen in Saccharomyces cerevisiae reveals a critical role for the mitochondria in the toxicity of a trichothecene mycotoxin.Proc Natl Acad Sci U S A. 2009 Dec 22;106(51):21883-8. doi: 10.1073/pnas.0909777106. Epub 2009 Dec 9. Proc Natl Acad Sci U S A. 2009. PMID: 20007368 Free PMC article.
-
ZNF9 activation of IRES-mediated translation of the human ODC mRNA is decreased in myotonic dystrophy type 2.PLoS One. 2010 Feb 18;5(2):e9301. doi: 10.1371/journal.pone.0009301. PLoS One. 2010. PMID: 20174632 Free PMC article.
References
-
- Albrecht M., Lengauer T. Novel Sm-like proteins with long C-terminal tails and associated methyltransferases. FEBS Lett. 2004;569:18–26. - PubMed
-
- Altschul S.F., Gish W., Miller W., Myers E.W., Lipman D.J. Basic local alignment search tool. J. Mol. Biol. 1990;215:403–410. - PubMed
-
- Anand M., Chakraburtty K., Marton M.J., Hinnebusch A.G., Kinzy T.G. Functional interactions between yeast translation eukaryotic elongation factor (eEF) 1A and eEF3. J. Biol. Chem. 2003;278:6985–6991. - PubMed
-
- Barbacid M., Vazquez D. (3H)anisomycin binding to eukaryotic ribosomes. J. Mol. Biol. 1974;84:603–623. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- N01AI40079/AI/NIAID NIH HHS/United States
- CA098131/CA/NCI NIH HHS/United States
- P01 HL068744/HL/NHLBI NIH HHS/United States
- P50 CA098131/CA/NCI NIH HHS/United States
- T32 AI49824/AI/NIAID NIH HHS/United States
- T32 AI049824/AI/NIAID NIH HHS/United States
- GM64779/GM/NIGMS NIH HHS/United States
- R01 GM064779/GM/NIGMS NIH HHS/United States
- R01 GM068900/GM/NIGMS NIH HHS/United States
- ES11993/ES/NIEHS NIH HHS/United States
- GM68900/GM/NIGMS NIH HHS/United States
- HL68744/HL/NHLBI NIH HHS/United States
- R01 ES011993/ES/NIEHS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases