

Ripening in the tomato Green-ripe mutant is inhibited by ectopic expression of a protein that disrupts ethylene signaling
- PMID: 16682641
- PMCID: PMC1458509
- DOI: 10.1073/pnas.0602319103
Ripening in the tomato Green-ripe mutant is inhibited by ectopic expression of a protein that disrupts ethylene signaling
Abstract
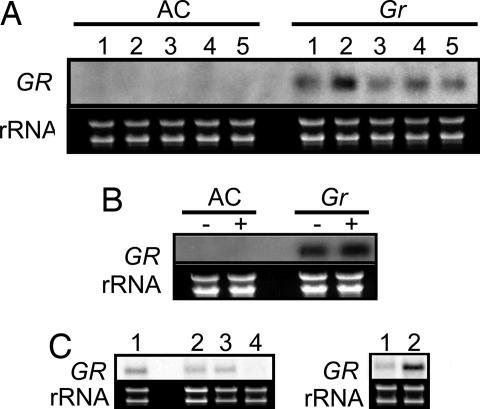
To achieve full ripening, climacteric fruits, such as tomato require synthesis, perception and signal transduction of the plant hormone ethylene. The nonripening phenotype of the dominant Green-ripe (Gr) and Never-ripe 2 (Nr-2) mutants of tomato is the result of reduced ethylene responsiveness in fruit tissues. In addition, a subset of ethylene responses associated with floral senescence, abscission, and root elongation are also impacted in mutant plants, but to a lesser extent. Using positional cloning, we have identified an identical 334-bp deletion in a gene of unknown biochemical function at the Gr/Nr-2 locus. Consistent with a dominant gain of function mutation, this deletion causes ectopic expression of Gr/Nr-2, which in turn leads to ripening inhibition. A CaMV35::GR transgene recreates the Gr/Nr-2 mutant phenotype but does not lead to a global reduction in ethylene responsiveness, suggesting tissue-specific modulation of ethylene responses in tomato. Gr/Nr-2 encodes an evolutionary conserved protein of unknown biochemical function that we associate here with ethylene signaling. Because Gr/Nr-2 has no sequence homology with the previously described Nr (Never-ripe) ethylene receptor of tomato we now refer to this gene only as GR. Identification of GR expands the current repertoire of ethylene signaling components in plants and provides a tool for further elucidation of ethylene response mechanisms and for controlling ethylene signal specificity in crop plants.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
Comment in
-
Highly conserved proteins that modify plant ethylene responses.Proc Natl Acad Sci U S A. 2006 May 16;103(20):7537-8. doi: 10.1073/pnas.0602599103. Epub 2006 May 8. Proc Natl Acad Sci U S A. 2006. PMID: 16682619 Free PMC article. No abstract available.
Similar articles
-
Ethylene insensitivity conferred by the Green-ripe and Never-ripe 2 ripening mutants of tomato.Plant Physiol. 2005 May;138(1):267-75. doi: 10.1104/pp.104.057745. Epub 2005 Apr 15. Plant Physiol. 2005. PMID: 15834010 Free PMC article.
-
Differential control of ethylene responses by GREEN-RIPE and GREEN-RIPE LIKE1 provides evidence for distinct ethylene signaling modules in tomato.Plant Physiol. 2012 Dec;160(4):1968-84. doi: 10.1104/pp.112.205476. Epub 2012 Oct 5. Plant Physiol. 2012. PMID: 23043080 Free PMC article.
-
Antisense inhibition of the Nr gene restores normal ripening to the tomato Never-ripe mutant, consistent with the ethylene receptor-inhibition model.Plant Physiol. 2000 Nov;124(3):1079-86. doi: 10.1104/pp.124.3.1079. Plant Physiol. 2000. PMID: 11080285 Free PMC article.
-
Ethylene Control of Fruit Ripening: Revisiting the Complex Network of Transcriptional Regulation.Plant Physiol. 2015 Dec;169(4):2380-90. doi: 10.1104/pp.15.01361. Epub 2015 Oct 28. Plant Physiol. 2015. PMID: 26511917 Free PMC article. Review.
-
Ethylene receptors and related proteins in climacteric and non-climacteric fruits.Plant Sci. 2018 Nov;276:63-72. doi: 10.1016/j.plantsci.2018.07.012. Epub 2018 Aug 10. Plant Sci. 2018. PMID: 30348329 Review.
Cited by
-
Undergraduates achieve learning gains in plant genetics through peer teaching of secondary students.CBE Life Sci Educ. 2014 Winter;13(4):641-52. doi: 10.1187/cbe.14-01-0007. CBE Life Sci Educ. 2014. PMID: 25452487 Free PMC article.
-
Genetic regulation and structural changes during tomato fruit development and ripening.Front Plant Sci. 2014 Apr 23;5:124. doi: 10.3389/fpls.2014.00124. eCollection 2014. Front Plant Sci. 2014. PMID: 24795731 Free PMC article. Review.
-
Role of the Tomato Non-Ripening Mutation in Regulating Fruit Quality Elucidated Using iTRAQ Protein Profile Analysis.PLoS One. 2016 Oct 12;11(10):e0164335. doi: 10.1371/journal.pone.0164335. eCollection 2016. PLoS One. 2016. PMID: 27732677 Free PMC article.
-
How ethylene works in the reproductive organs of higher plants: a signaling update from the third millennium.Plant Signal Behav. 2006 Sep;1(5):231-42. doi: 10.4161/psb.1.5.3389. Plant Signal Behav. 2006. PMID: 19516984 Free PMC article.
-
Malate plays a crucial role in starch metabolism, ripening, and soluble solid content of tomato fruit and affects postharvest softening.Plant Cell. 2011 Jan;23(1):162-84. doi: 10.1105/tpc.109.072231. Epub 2011 Jan 14. Plant Cell. 2011. PMID: 21239646 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
