

A leaderless mRNA can bind to mammalian 80S ribosomes and direct polypeptide synthesis in the absence of translation initiation factors
- PMID: 16581790
- PMCID: PMC1446950
- DOI: 10.1128/MCB.26.8.3164-3169.2006
A leaderless mRNA can bind to mammalian 80S ribosomes and direct polypeptide synthesis in the absence of translation initiation factors
Abstract
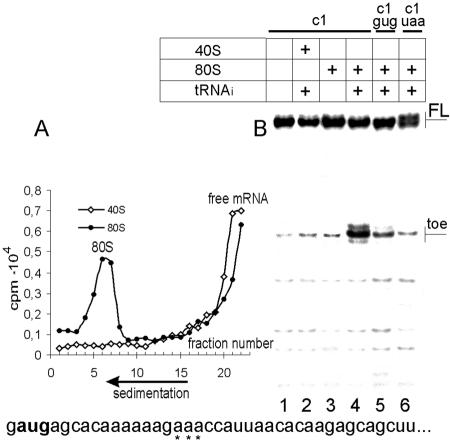
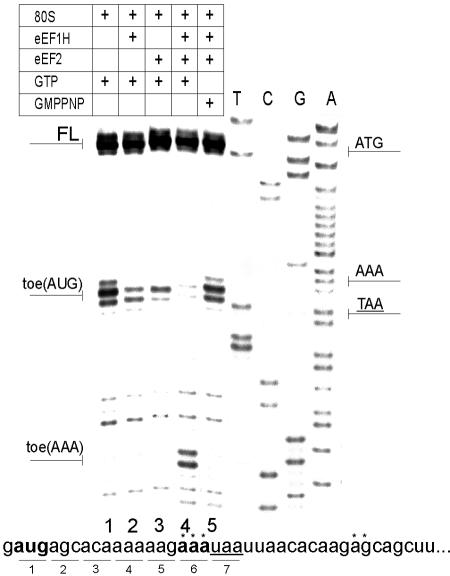
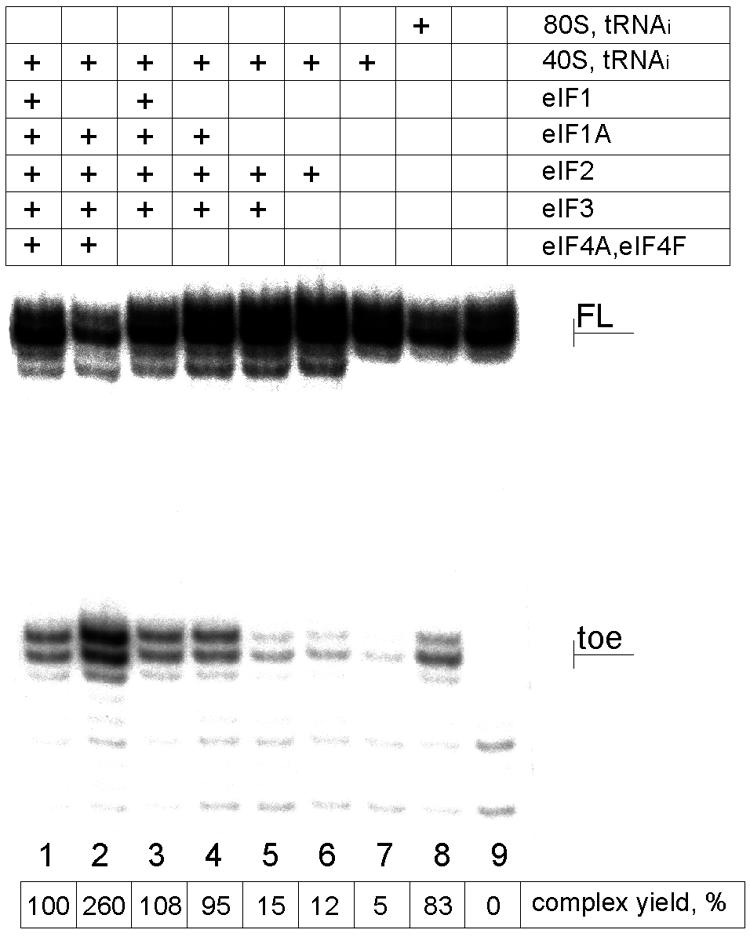
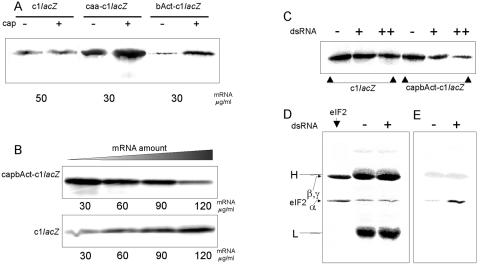
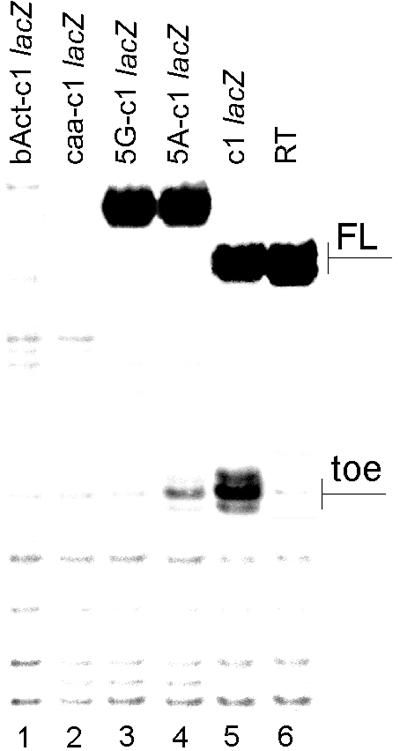
Translation initiation in eukaryotic cells is known to be a complex multistep process which involves numerous protein factors. Here we demonstrate that leaderless mRNAs with initiator Met-tRNA can bind directly to 80S mammalian ribosomes in the absence of initiation factors and that the complexes thus formed are fully competent for the subsequent steps of polypeptide synthesis. We show that the canonical 48S pathway of eukaryotic translation initiation has no obvious advantage over the 80S pathway of translation initiation on leaderless mRNAs and suggest that, in the presence of competing mRNAs containing a leader, the latter mechanism will be preferred. The direct binding of the leaderless mRNA to the 80S ribosome was precluded when such an mRNA was supplied with a 5' leader, irrespective of whether it was in a totally single-stranded conformation or was prone to base pairing. The striking similarity between the mechanisms of binding of leaderless mRNAs with mammalian 80S or bacterial 70S ribosomes gives support to the idea that the alternative mode of translation initiation used by leaderless mRNAs represents a relic from early steps in the evolution of the translation apparatus.
Figures
Similar articles
-
Selective stimulation of translation of leaderless mRNA by initiation factor 2: evolutionary implications for translation.EMBO J. 2000 Aug 1;19(15):4101-10. doi: 10.1093/emboj/19.15.4101. EMBO J. 2000. PMID: 10921890 Free PMC article.
-
Evolution and the universality of the mechanism of initiation of protein synthesis.Gene. 2009 Mar 1;432(1-2):1-6. doi: 10.1016/j.gene.2008.11.001. Epub 2008 Nov 8. Gene. 2009. PMID: 19056476 Review.
-
Translation initiation with 70S ribosomes: an alternative pathway for leaderless mRNAs.Nucleic Acids Res. 2004 Jun 23;32(11):3354-63. doi: 10.1093/nar/gkh663. Print 2004. Nucleic Acids Res. 2004. PMID: 15215335 Free PMC article.
-
Translation initiation factor 3 antagonizes authentic start codon selection on leaderless mRNAs.Mol Microbiol. 1999 Jan;31(1):67-77. doi: 10.1046/j.1365-2958.1999.01147.x. Mol Microbiol. 1999. PMID: 9987111
-
Translation initiation by factor-independent binding of eukaryotic ribosomes to internal ribosomal entry sites.C R Biol. 2005 Jul;328(7):589-605. doi: 10.1016/j.crvi.2005.02.004. C R Biol. 2005. PMID: 15992743 Review.
Cited by
-
Experimental characterization of Cis-acting elements important for translation and transcription in halophilic archaea.PLoS Genet. 2007 Dec;3(12):e229. doi: 10.1371/journal.pgen.0030229. PLoS Genet. 2007. PMID: 18159946 Free PMC article.
-
Activation of the antiviral factor RNase L triggers translation of non-coding mRNA sequences.Nucleic Acids Res. 2021 Jun 21;49(11):6007-6026. doi: 10.1093/nar/gkab036. Nucleic Acids Res. 2021. PMID: 33556964 Free PMC article.
-
Comparison of mRNA features affecting translation initiation and reinitiation.Nucleic Acids Res. 2013 Jan 7;41(1):474-86. doi: 10.1093/nar/gks989. Epub 2012 Oct 23. Nucleic Acids Res. 2013. PMID: 23093605 Free PMC article.
-
[Nonspecific and specific interaction of Y-box binding protein 1 (YB-1) with mRNA and posttranscriptional regulation of protein synthesis in animal cells].Mol Biol (Mosk). 2006 Jul-Aug;40(4):620-33. doi: 10.1134/s0026893306040145. Mol Biol (Mosk). 2006. PMID: 16913221 Review. Russian.
-
Ribosomes bind leaderless mRNA in Escherichia coli through recognition of their 5'-terminal AUG.RNA. 2008 Oct;14(10):2159-69. doi: 10.1261/rna.1089208. Epub 2008 Aug 28. RNA. 2008. PMID: 18755843 Free PMC article.
References
-
- Battiste, J. L., T. V. Pestova, C. U. Hellen, and G. Wagner. 2000. The eIF1A solution structure reveals a large RNA-binding surface important for scanning function. Mol. Cell 5:109-119. - PubMed
-
- Dmitriev, S. E., A. V. Pisarev, M. P. Rubtsova, Y. E. Dunaevsky, and I. N. Shatsky. 2003. Conversion of 48S translation preinitiation complexes into 80S initiation complexes as revealed by toeprinting. FEBS Lett. 533:99-104. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous