

Inhibition of ADP/ATP exchange in receptor-interacting protein-mediated necrosis
- PMID: 16507998
- PMCID: PMC1430284
- DOI: 10.1128/MCB.26.6.2215-2225.2006
Inhibition of ADP/ATP exchange in receptor-interacting protein-mediated necrosis
Abstract
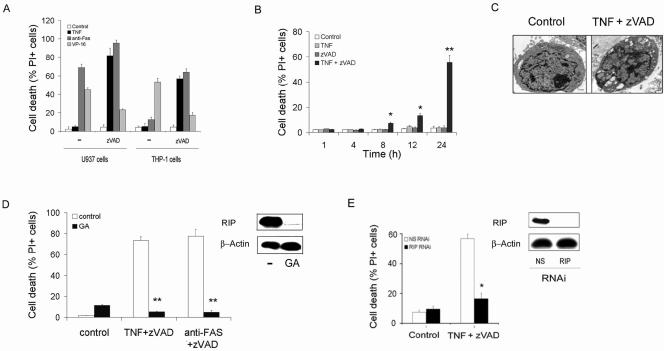
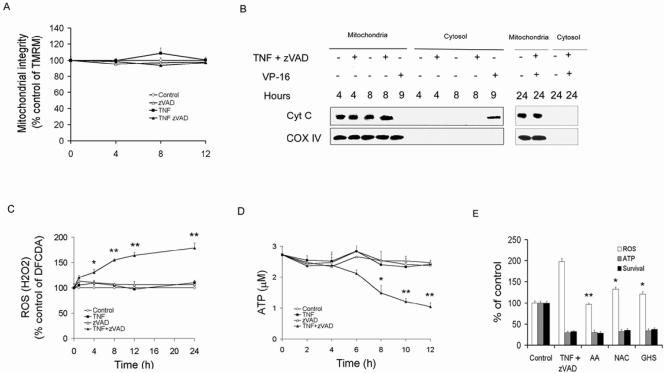
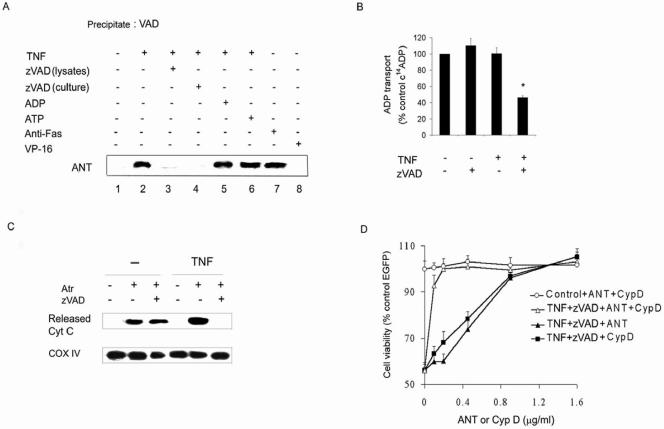
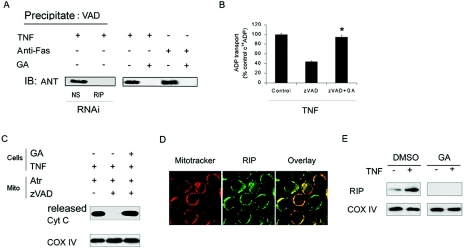
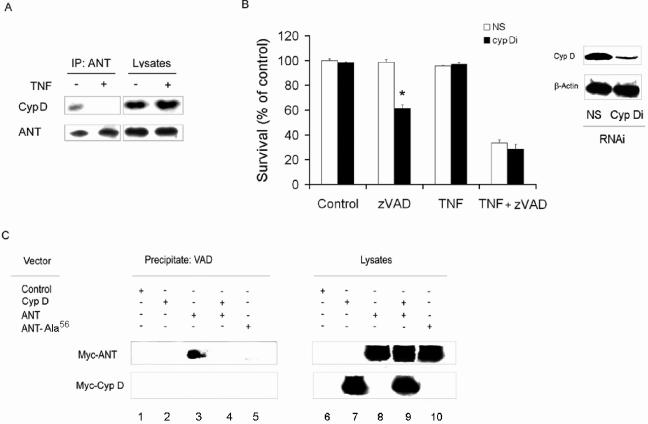
Receptor-interacting protein (RIP) has been implicated in the induction of death receptor-mediated, nonapoptotic cell death. However, the mechanisms remain to be elucidated. Here we show that tumor necrosis factor alpha induced RIP-dependent inhibition of adenine nucleotide translocase (ANT)-conducted transport of ADP into mitochondria, which resulted in reduced ATP and necrotic cell death. The inhibition of ADP/ATP exchange coincided with the loss of interaction between ANT and cyclophilin D and the inability of ANT to adopt the cytosolic conformational state, which prevented cytochrome c release. Neither overexpression of Bcl-xL nor inhibition of reactive oxygen species prevented necrosis. In contrast, the ectopic expression of ANT or cyclophilin D was effective at preventing cell death. These observations demonstrate a novel mechanism initiated through death receptor ligation and mediated by RIP that results in the suppression of ANT activity and necrosis.
Figures
Similar articles
-
Bcl-2 and Bax modulate adenine nucleotide translocase activity.Cancer Res. 2003 Jan 15;63(2):541-6. Cancer Res. 2003. PMID: 12543814
-
Role of critical thiol groups on the matrix surface of the adenine nucleotide translocase in the mechanism of the mitochondrial permeability transition pore.Biochem J. 2002 Oct 15;367(Pt 2):541-8. doi: 10.1042/BJ20011672. Biochem J. 2002. PMID: 12149099 Free PMC article.
-
Caspase inhibition switches the mode of cell death induced by cyanide by enhancing reactive oxygen species generation and PARP-1 activation.Toxicol Appl Pharmacol. 2004 Mar 1;195(2):194-202. doi: 10.1016/j.taap.2003.11.012. Toxicol Appl Pharmacol. 2004. PMID: 14998685
-
Caspase inhibitors promote alternative cell death pathways.Sci STKE. 2006 Oct 24;2006(358):pe44. doi: 10.1126/stke.3582006pe44. Sci STKE. 2006. PMID: 17062895 Review.
-
The adenine nucleotide translocase: a central component of the mitochondrial permeability transition pore and key player in cell death.Curr Med Chem. 2003 Aug;10(16):1507-25. doi: 10.2174/0929867033457278. Curr Med Chem. 2003. PMID: 12871123 Review.
Cited by
-
Pyroptosis in health and disease: mechanisms, regulation and clinical perspective.Signal Transduct Target Ther. 2024 Sep 20;9(1):245. doi: 10.1038/s41392-024-01958-2. Signal Transduct Target Ther. 2024. PMID: 39300122 Free PMC article. Review.
-
Adenine nucleotide (ADP/ATP) translocase 3 participates in the tumor necrosis factor induced apoptosis of MCF-7 cells.Mol Biol Cell. 2007 Nov;18(11):4681-9. doi: 10.1091/mbc.e06-12-1161. Epub 2007 Sep 12. Mol Biol Cell. 2007. PMID: 17855512 Free PMC article.
-
Neuronal cell death in neonatal hypoxia-ischemia.Ann Neurol. 2011 May;69(5):743-58. doi: 10.1002/ana.22419. Ann Neurol. 2011. PMID: 21520238 Free PMC article.
-
RIP kinase-mediated necrosis as an alternative mechanisms of photoreceptor death.Oncotarget. 2011 Jun;2(6):497-509. doi: 10.18632/oncotarget.286. Oncotarget. 2011. PMID: 21670490 Free PMC article. Review.
-
Cell death in pancreatic cancer: from pathogenesis to therapy.Nat Rev Gastroenterol Hepatol. 2021 Nov;18(11):804-823. doi: 10.1038/s41575-021-00486-6. Epub 2021 Jul 30. Nat Rev Gastroenterol Hepatol. 2021. PMID: 34331036 Review.
References
-
- Ahmad, M., S. M. Srinivasula, L. Wang, R. V. Talanian, G. Litwack, T. Fernandes-Alnemri, and E. S. Alnemri. 1997. CRADD, a novel human apoptotic adaptor molecule for caspase-2, and FasL/tumor necrosis factor receptor-interacting protein RIP. Cancer Res. 57:615-619. - PubMed
-
- Baines, C. P., R. A. Kaiser, N. H. Purcell, N. S. Blair, H. Osinska, M. A. Hambleton, E. W. Brunskill, M. R. Sayen, R. A. Gottlieb, G. W. Dorn, J. Robbins, and J. D. Molkentin. 2005. Loss of cyclophilin D reveals a critical role for mitochondrial permeability transition in cell death. Nature 434:658-662. - PubMed
-
- Belzacq, A. S., H. L. Vieira, F. Verrier, G. Vandecasteele, I. Cohen, M. C. Prevost, E. Larquet, F. Pariselli, P. X. Petit, A. Kahn, R. Rizzuto, C. Brenner, and G. Kroemer. 2003. Bcl-2 and Bax modulate adenine nucleotide translocase activity. Cancer Res. 63:541-546. - PubMed
-
- Bottero, V., F. Rossi, M. Samson, M. Mari, P. Hofman, and J. F. Peyron. 2001. Ikappa b-alpha, the NF-kappa B inhibitory subunit, interacts with ANT, the mitochondrial ATP/ADP translocator. J. Biol. Chem. 276:21317-21324. - PubMed
-
- Castedo, M., K. Ferri, T. Roumier, D. Metivier, N. Zamzami, and G. Kroemer. 2002. Quantitation of mitochondrial alterations associated with apoptosis. J. Immunol. Methods 265:39-47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous