

Identification of Rhodospirillum rubrum GlnB variants that are altered in their ability to interact with different targets in response to nitrogen status signals
- PMID: 16484197
- PMCID: PMC1426566
- DOI: 10.1128/JB.188.5.1866-1874.2006
Identification of Rhodospirillum rubrum GlnB variants that are altered in their ability to interact with different targets in response to nitrogen status signals
Abstract
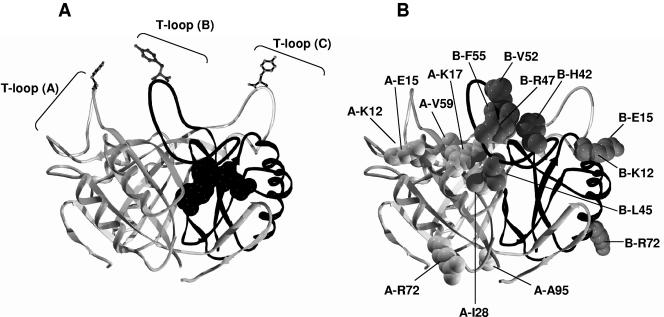
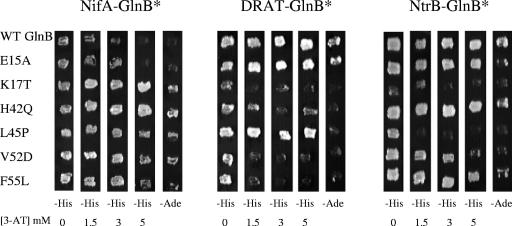
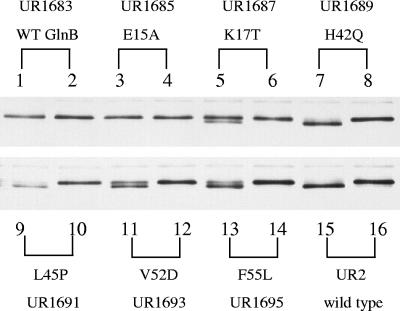
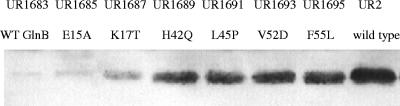
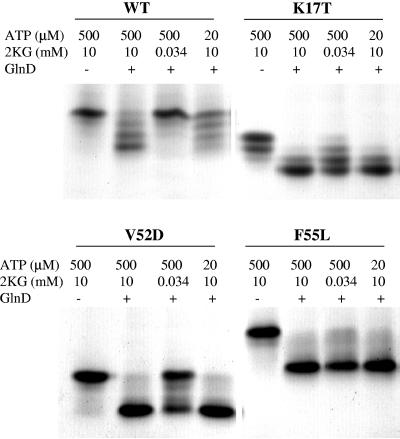
In Rhodospirillum rubrum, NifA, the transcriptional activator for the nif genes, is posttranslationally activated only by the uridylylated form of GlnB, one of three P(II) homologs in the organism. We have used the yeast two-hybrid system to detect variants of GlnB that interact better with NifA than does wild-type GlnB. When examined for physiological effects in R. rubrum, these GlnB* variants activated NifA in the presence of NH(4)(+), which normally blocks NifA activation completely, and in the absence of GlnD, whose uridylylation of GlnB is also normally essential for NifA activation. When these variants were tested in the two-hybrid system for their interaction with NtrB, a receptor that should interact with the nonuridylylated form of GlnB, they were uniformly weaker than wild-type GlnB in that interaction. When expressed in R. rubrum either as single-copy integrants or on multiple-copy plasmids, these variants were also dramatically altered in terms of their ability to regulate several other receptors involved in nitrogen metabolism, including GlnE, NtrB/NtrC, and DRAT (dinitrogenase reductase ADP-ribosyl transferase)-DRAG (dinitrogenase reductase-activating glycohydrolase). The consistent pattern throughout is that these GlnB variants partially mimic the uridylylated form of wild-type GlnB, even under nitrogen-excess conditions and in strains lacking GlnD. The results suggest that the role of uridylylation of GlnB is primarily to shift the equilibrium of GlnB from a "nitrogen-sufficient" form to a "nitrogen-deficient" form, each of which interacts with different but overlapping receptor proteins in the cell. These GlnB variants apparently shift that equilibrium through direct structural changes.
Figures
Similar articles
-
Mutagenesis and functional characterization of the glnB, glnA, and nifA genes from the photosynthetic bacterium Rhodospirillum rubrum.J Bacteriol. 2000 Feb;182(4):983-92. doi: 10.1128/JB.182.4.983-992.2000. J Bacteriol. 2000. PMID: 10648524 Free PMC article.
-
Effect of P(II) and its homolog GlnK on reversible ADP-ribosylation of dinitrogenase reductase by heterologous expression of the Rhodospirillum rubrum dinitrogenase reductase ADP-ribosyl transferase-dinitrogenase reductase-activating glycohydrolase regulatory system in Klebsiella pneumoniae.J Bacteriol. 2001 Mar;183(5):1610-20. doi: 10.1128/JB.183.5.1610-1620.2001. J Bacteriol. 2001. PMID: 11160092 Free PMC article.
-
Identification and functional characterization of NifA variants that are independent of GlnB activation in the photosynthetic bacterium Rhodospirillum rubrum.Microbiology (Reading). 2008 Sep;154(Pt 9):2689-2699. doi: 10.1099/mic.0.2008/019406-0. Microbiology (Reading). 2008. PMID: 18757802 Free PMC article.
-
Regulation of nitrogen fixation in Klebsiella pneumoniae and Azotobacter vinelandii: NifL, transducing two environmental signals to the nif transcriptional activator NifA.J Mol Microbiol Biotechnol. 2002 May;4(3):235-42. J Mol Microbiol Biotechnol. 2002. PMID: 11931553 Review.
-
Metabolic regulation of nitrogen fixation in Rhodospirillum rubrum.Biochem Soc Trans. 2006 Feb;34(Pt 1):160-1. doi: 10.1042/BST0340160. Biochem Soc Trans. 2006. PMID: 16417510 Review.
Cited by
-
The nitrogenase regulatory enzyme dinitrogenase reductase ADP-ribosyltransferase (DraT) is activated by direct interaction with the signal transduction protein GlnB.J Bacteriol. 2013 Jan;195(2):279-86. doi: 10.1128/JB.01517-12. Epub 2012 Nov 9. J Bacteriol. 2013. PMID: 23144248 Free PMC article.
-
Mutagenesis and functional characterization of the four domains of GlnD, a bifunctional nitrogen sensor protein.J Bacteriol. 2010 Jun;192(11):2711-21. doi: 10.1128/JB.01674-09. Epub 2010 Apr 2. J Bacteriol. 2010. PMID: 20363937 Free PMC article.
-
In vitro studies of the uridylylation of the three PII protein paralogs from Rhodospirillum rubrum: the transferase activity of R. rubrum GlnD is regulated by alpha-ketoglutarate and divalent cations but not by glutamine.J Bacteriol. 2007 May;189(9):3471-8. doi: 10.1128/JB.01704-06. Epub 2007 Mar 2. J Bacteriol. 2007. PMID: 17337583 Free PMC article.
-
Effect of perturbation of ATP level on the activity and regulation of nitrogenase in Rhodospirillum rubrum.J Bacteriol. 2009 Sep;191(17):5526-37. doi: 10.1128/JB.00585-09. Epub 2009 Jun 19. J Bacteriol. 2009. PMID: 19542280 Free PMC article.
-
In vitro interactions between the PII proteins and the nitrogenase regulatory enzymes dinitrogenase reductase ADP-ribosyltransferase (DraT) and dinitrogenase reductase-activating glycohydrolase (DraG) in Azospirillum brasilense.J Biol Chem. 2009 Mar 13;284(11):6674-82. doi: 10.1074/jbc.M807378200. Epub 2009 Jan 8. J Biol Chem. 2009. PMID: 19131333 Free PMC article.
References
-
- Atkinson, M. R., E. S. Kamberov, R. L. Weiss, and A. J. Ninfa. 1994. Reversible uridylylation of the Escherichia coli PII signal transduction protein regulates its ability to stimulate the dephosphorylation of the transcription factor nitrogen regulator I (NRI or NtrC). J. Biol. Chem. 269:28288-28293. - PubMed
-
- Atkinson, M. R., and A. J. Ninfa. 1998. Role of the GlnK signal transduction protein in the regulation of nitrogen assimilation in Escherichia coli. Mol. Microbiol. 29:431-447. - PubMed
-
- Benelli, E. M., M. Buck, I. Polikarpov, E. M. Souza, L. M. Cruz, and F. O. Pedrosa. 2002. Herbaspirillum seropedicae signal transduction protein PII is structurally similar to the enteric GlnK. Eur. J. Biochem. 269:3296-3303. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
