

The Nfs1 interacting protein Isd11 has an essential role in Fe/S cluster biogenesis in mitochondria
- PMID: 16341090
- PMCID: PMC1356348
- DOI: 10.1038/sj.emboj.7600905
The Nfs1 interacting protein Isd11 has an essential role in Fe/S cluster biogenesis in mitochondria
Abstract
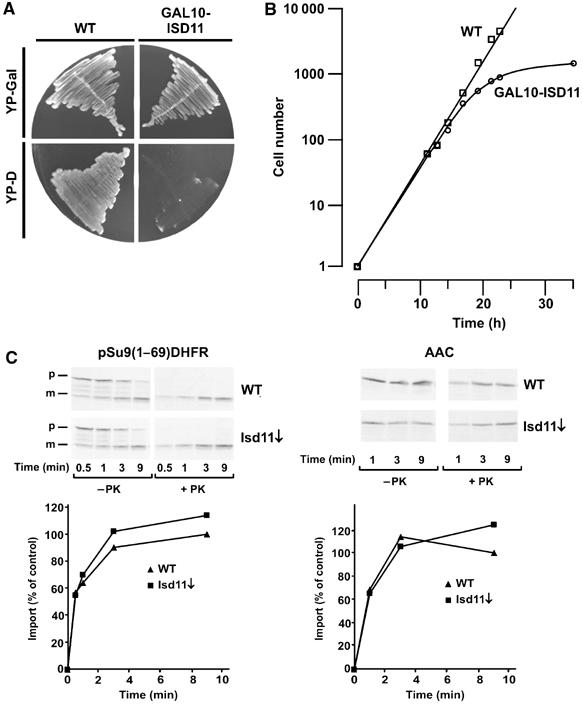
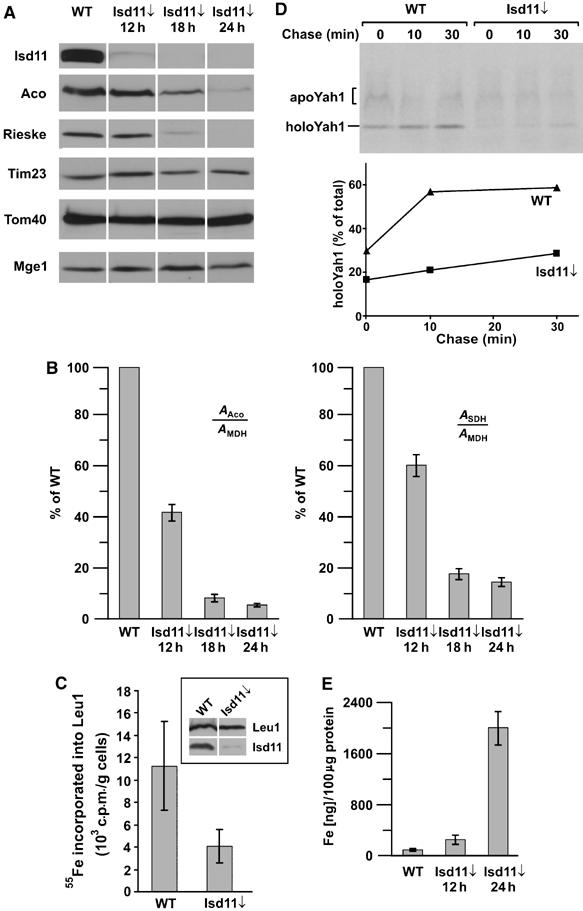
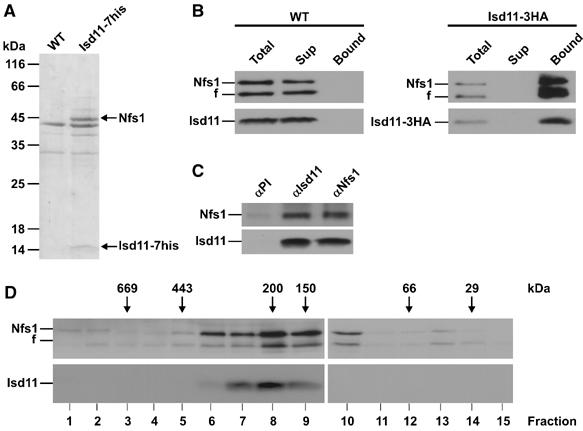
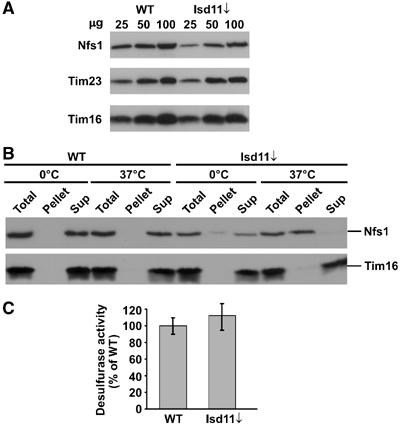
Formation of iron/sulfur (Fe/S) clusters, protein translocation and protein folding are essential processes in the mitochondria of Saccharomyces cerevisiae. In a systematic approach to characterize essential proteins involved in these processes, we identified a novel essential protein of the mitochondrial matrix, which is highly conserved from yeast to human and which we termed Isd11. Depletion of Isd11 caused a strong reduction in the levels of the Fe/S proteins aconitase and the Rieske protein, and a massive decrease in the enzymatic activities of aconitase and succinate dehydrogenase. Incorporation of iron into the Fe/S protein Leu1 and formation of the Fe/S cluster containing holoform of the mitochondrial ferredoxin Yah1 were inhibited in the absence of Isd11. This strongly suggests that Isd11 is required for the assembly of Fe/S proteins. We show that Isd11 forms a stable complex with Nfs1, the cysteine desulfurase of the mitochondrial machinery for Fe/S cluster assembly. In the absence of Isd11, Nfs1 is prone to aggregation. We propose that Isd11 acts together with Nfs1 in an early step in the biogenesis of Fe/S proteins.
Figures

Similar articles
-
Essential role of Isd11 in mitochondrial iron-sulfur cluster synthesis on Isu scaffold proteins.EMBO J. 2006 Jan 11;25(1):184-95. doi: 10.1038/sj.emboj.7600906. Epub 2005 Dec 8. EMBO J. 2006. PMID: 16341089 Free PMC article.
-
Mitochondrial frataxin interacts with ISD11 of the NFS1/ISCU complex and multiple mitochondrial chaperones.Hum Mol Genet. 2007 Apr 15;16(8):929-41. doi: 10.1093/hmg/ddm038. Epub 2007 Mar 1. Hum Mol Genet. 2007. PMID: 17331979
-
Predictive reconstruction of the mitochondrial iron-sulfur cluster assembly metabolism. II. Role of glutaredoxin Grx5.Proteins. 2004 Nov 15;57(3):481-92. doi: 10.1002/prot.20228. Proteins. 2004. PMID: 15382238
-
Iron-sulfur cluster biosynthesis in bacteria: Mechanisms of cluster assembly and transfer.Arch Biochem Biophys. 2008 Jun 15;474(2):226-37. doi: 10.1016/j.abb.2007.12.014. Epub 2007 Dec 28. Arch Biochem Biophys. 2008. PMID: 18191630 Review.
-
The role of mitochondria in cellular iron-sulfur protein biogenesis and iron metabolism.Biochim Biophys Acta. 2012 Sep;1823(9):1491-508. doi: 10.1016/j.bbamcr.2012.05.009. Epub 2012 May 15. Biochim Biophys Acta. 2012. PMID: 22609301 Review.
Cited by
-
Biogenesis of iron-sulfur clusters in mammalian cells: new insights and relevance to human disease.Dis Model Mech. 2012 Mar;5(2):155-64. doi: 10.1242/dmm.009019. Dis Model Mech. 2012. PMID: 22382365 Free PMC article. Review.
-
Structural diversity of cysteine desulfurases involved in iron-sulfur cluster biosynthesis.Biophys Physicobiol. 2022 Feb 8;19:1-18. doi: 10.2142/biophysico.bppb-v19.0001. eCollection 2022. Biophys Physicobiol. 2022. PMID: 35377584 Free PMC article.
-
Isd11p protein activates the mitochondrial cysteine desulfurase Nfs1p protein.J Biol Chem. 2011 Nov 4;286(44):38242-38252. doi: 10.1074/jbc.M111.288522. Epub 2011 Sep 9. J Biol Chem. 2011. Retraction in: J Biol Chem. 2012 Jun 22;287(26):22152. doi: 10.1074/jbc.a111.288522 PMID: 21908622 Free PMC article. Retracted.
-
Respiratory chain supercomplexes associate with the cysteine desulfurase complex of the iron-sulfur cluster assembly machinery.Mol Biol Cell. 2018 Apr 1;29(7):776-785. doi: 10.1091/mbc.E17-09-0555. Mol Biol Cell. 2018. PMID: 29386296 Free PMC article.
-
Iron‑sulfur clusters in viral proteins: Exploring their elusive nature, roles and new avenues for targeting infections.Biochim Biophys Acta Mol Cell Res. 2024 Jun;1871(5):119723. doi: 10.1016/j.bbamcr.2024.119723. Epub 2024 Apr 8. Biochim Biophys Acta Mol Cell Res. 2024. PMID: 38599324 Review.
References
-
- Babcock M, de Silva D, Oaks R, Davis-Kaplan S, Jiralerspong S, Montermini L, Pandolfo M, Kaplan J (1997) Regulation of mitochondrial iron accumulation by Yfh1p, a putative homolog of frataxin. Science 276: 1709–1712 - PubMed
-
- Cheng MY, Hartl FU, Martin J, Pollock RA, Kalousek F, Neupert W, Hallberg EM, Hallberg RL, Horwich AL (1989) Mitochondrial heat-shock protein hsp60 is essential for assembly of proteins imported into yeast mitochondria. Nature 337: 620–625 - PubMed
-
- Craig EA, Voisine C, Schilke B (1999) Mitochondrial iron metabolism in the yeast Saccharomyces cerevisiae. Biol Chem 380: 1167–1173 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
