

Review
doi: 10.1083/jcb.200508051.
Signal-mediated export of proteins from the malaria parasite to the host erythrocyte
Affiliations
- PMID: 16301328
- PMCID: PMC2171567
- DOI: 10.1083/jcb.200508051
Item in Clipboard
Review
Signal-mediated export of proteins from the malaria parasite to the host erythrocyte
J Cell Biol.
.
Abstract
Intracellular parasites from the genus Plasmodium reside and multiply in a variety of cells during their development. After invasion of human erythrocytes, asexual stages from the most virulent malaria parasite, P. falciparum, drastically change their host cell and export remodelling and virulence proteins. Recent data demonstrate that a specific NH(2)-terminal signal conserved across the genus Plasmodium plays a central role in this export process.
Figures
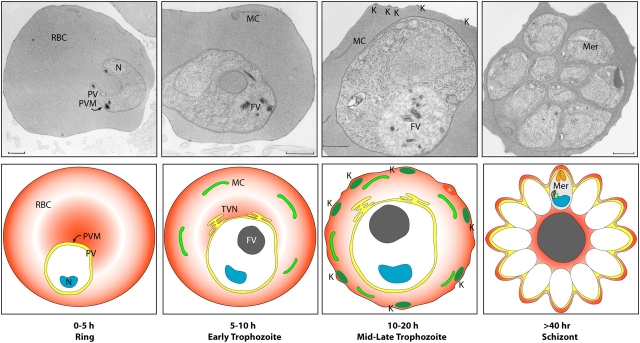
Asexual development of malaria parasites in the iRBC. 0–5 h: after invasion, parasites become surrounded by a parasitophorous vacuole (PV) and are visible as ring-like structures within the red blood cell (RBC; ring stage). 5–10 h: the ring-stage parasite induces membranous extensions of the parasitophorous vacuole membrane (PVM) into the host cell, which forms a tubulovesicular network (TVN) and RBC membrane–tethered Maurer's clefts (MCs). The products of hemoglobin digestion become visible with the deposition of hemozoin crystals in the food vacuole (FV). 10–20 h: as the parasite progresses to the trophozoite stage, it continues to increase its volume considerably and induces knob (K) formation on the iRBC surface. >40 h: after invasion, the parasite starts several rounds of asexual division, resulting in 16–32 daughter merozoites, which are still surrounded by the PVM until they burst out and invade uninfected RBCs. Mer, merozoite; N, nucleus. Bars, 0.5 μm.
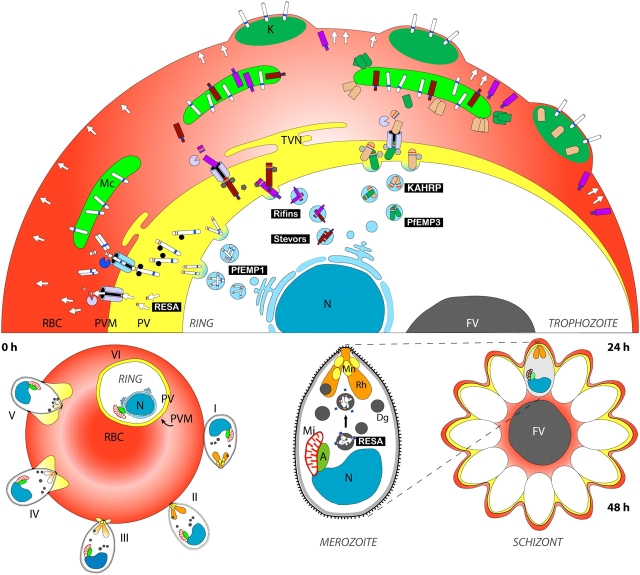
A model for signal-mediated protein export during the asexual cycle. Approximately 400 parasite proteins have a predicted PEXEL motif that is expected to direct export to the RBC cytosol. Many of these may play roles in trafficking of PfEMP1 to the RBC surface or modifying the RBC membrane in other ways to suit the parasite's needs. Plasmodium prepares for the invasion and remodelling process of a new host cell in late asexual stages (schizont stage) by transporting crucial molecules to the three prototypic apicomplexan organelles (the microneme [Mn], the rhoptry [Rh], and the dense granule [Dg]) located close to the apical end of the parasite. While microneme proteins are mainly involved in initiating invasion (stages I–III), rhoptry and dense granule proteins are implicated in establishment of the parasite in the newly invaded host cell (stages III–VI). RESA (arrows), for example, is targeted to the dense granules in late schizogony, released into the parasitophorous vacuole (PV) after invasion, and subsequently exported into the red blood cell (RBC), where it associates with the red cell cytoskeleton to stabilize the newly invaded RBC membrane. The presence of a PEXEL motif in RESA implies the establishment of the corresponding translocation machinery very early after formation of the PVM. This allows the virulence factor PfEMP1 to be translocated already in early ring stages. It then appears to be transported as a soluble complex and inserted into the Maurer's cleft (MC) membrane. In trophozoite stages, knob components such as KAHRP and PfEMP3 are exported and travel as large complexes through the RBC cytosol until they transiently associate with components on the MC. Further transport of KAHRP, PfEMP3, and PfEMP1 leads to knob formation and surface exposition of PfEMP1. It is not known whether Rifin and Stevor proteins are also trafficked as soluble complexes in the RBC cytosol or whether there is vesicular transport between the PVM, MCs, and possibly the RBC membrane. Mi, mitochondrion; A, apicoplast. Other abbreviations are the same as those in Fig. 1.
Similar articles
-
The role of the Maurer's clefts in protein transport in Plasmodium falciparum.Trends Parasitol. 2009 Jun;25(6):277-84. doi: 10.1016/j.pt.2009.03.009. Epub 2009 May 11. Trends Parasitol. 2009. PMID: 19442584 Review.
-
Targeting malaria parasite proteins to the erythrocyte.Trends Parasitol. 2005 Sep;21(9):399-402. doi: 10.1016/j.pt.2005.07.006. Trends Parasitol. 2005. PMID: 16046185 Review.
-
Pexel/VTS: a protein-export motif in erythrocytes infected with malaria parasites.Trends Parasitol. 2005 Sep;21(9):396-9. doi: 10.1016/j.pt.2005.07.004. Trends Parasitol. 2005. PMID: 16046186 Review.
-
Targeting malaria virulence and remodeling proteins to the host erythrocyte.Science. 2004 Dec 10;306(5703):1930-3. doi: 10.1126/science.1102452. Science. 2004. PMID: 15591202
-
Protein targeting from malaria parasites to host erythrocytes.Traffic. 2005 Aug;6(8):706-9. doi: 10.1111/j.1600-0854.2005.00310.x. Traffic. 2005. PMID: 15998325 Review.
Cited by
-
Spatial association with PTEX complexes defines regions for effector export into Plasmodium falciparum-infected erythrocytes.Nat Commun. 2013;4:1415. doi: 10.1038/ncomms2449. Nat Commun. 2013. PMID: 23361006 Free PMC article.
-
Malaria parasite proteins that remodel the host erythrocyte.Nat Rev Microbiol. 2009 May;7(5):341-54. doi: 10.1038/nrmicro2110. Nat Rev Microbiol. 2009. PMID: 19369950 Review.
-
Inhibition of protein N-myristoylation blocks Plasmodium falciparum intraerythrocytic development, egress and invasion.PLoS Biol. 2021 Oct 25;19(10):e3001408. doi: 10.1371/journal.pbio.3001408. eCollection 2021 Oct. PLoS Biol. 2021. PMID: 34695132 Free PMC article.
-
Analysis of subtelomeric virulence gene families in Plasmodium falciparum by comparative transcriptional profiling.Mol Microbiol. 2012 Apr;84(2):243-59. doi: 10.1111/j.1365-2958.2012.08019.x. Epub 2012 Mar 22. Mol Microbiol. 2012. PMID: 22435676 Free PMC article.
-
Maurer's clefts of Plasmodium falciparum are secretory organelles that concentrate virulence protein reporters for delivery to the host erythrocyte.Blood. 2008 Feb 15;111(4):2418-26. doi: 10.1182/blood-2007-09-115279. Epub 2007 Dec 5. Blood. 2008. PMID: 18057226 Free PMC article.
References
-
- Ansorge, I., K. Paprotka, S. Bhakdi, and K. Lingelbach. 1997. Permeabilization of the erythrocyte membrane with streptolysin O allows access to the vacuolar membrane of Plasmodium falciparum and a molecular analysis of membrane topology. Mol. Biochem. Parasitol. 84:259–261. - PubMed
-
- Atkinson, C.T., and M. Aikawa. 1990. Ultrastructure of malaria-infected erythrocytes. Blood Cells. 16:351–368. - PubMed
-
- Atkinson, C.T., M. Aikawa, G. Perry, T. Fujino, V. Bennett, E.A. Davidson, and R.J. Howard. 1988. Ultrastructural localization of erythrocyte cytoskeletal and integral membrane proteins in Plasmodium falciparum-infected erythrocytes. Eur. J. Cell Biol. 45:192–199. - PubMed
-
- Bannister, L.H., J.M. Hopkins, G. Margos, A.R. Dluzewski, and G.H. Mitchell. 2004. Three-dimensional ultrastructure of the ring stage of Plasmodium falciparum: evidence for export pathways. Microsc. Microanal. 10:551–562. - PubMed
-
- Baruch, D.I., B.L. Pasloske, H.B. Singh, X. Bi, X.C. Ma, M. Feldman, T.F. Taraschi, and R.J. Howard. 1995. Cloning the P. falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell. 82:77–87. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
