

Heterogeneous nuclear ribonucleoprotein C1/C2, MeCP1, and SWI/SNF form a chromatin remodeling complex at the beta-globin locus control region
- PMID: 16217013
- PMCID: PMC1257739
- DOI: 10.1073/pnas.0507596102
Heterogeneous nuclear ribonucleoprotein C1/C2, MeCP1, and SWI/SNF form a chromatin remodeling complex at the beta-globin locus control region
Abstract
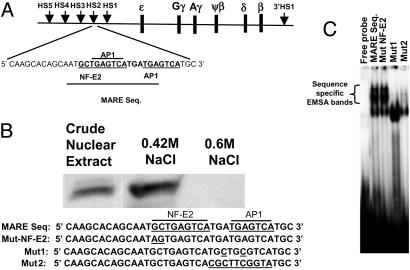
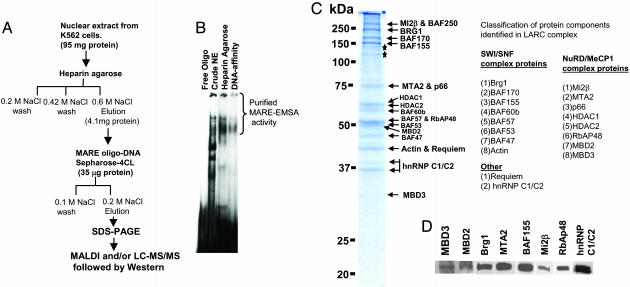
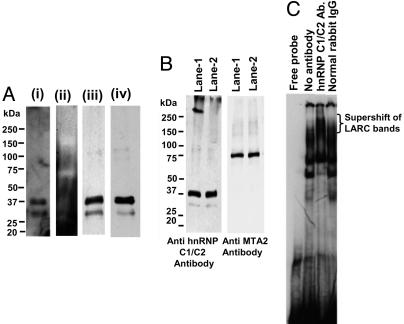
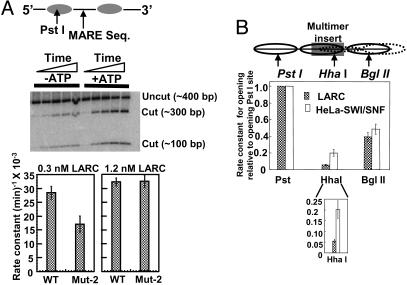
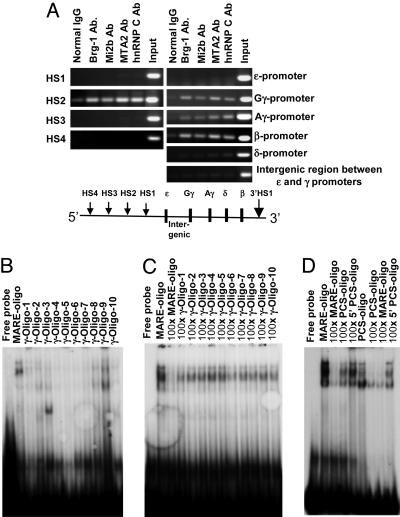
Locus control regions (LCRs) are regulatory DNA sequences that are situated many kilobases away from their cognate promoters. LCRs protect transgenes from position effect variegation and heterochromatinization and also promote copy-number dependence of the levels of transgene expression. In this work, we describe the biochemical purification of a previously undescribed LCR-associated remodeling complex (LARC) that consists of heterogeneous nuclear ribonucleoprotein C1/C2, nucleosome remodeling SWI/SNF, and nucleosome remodeling and deacetylating (NuRD)/MeCP1 as a single homogeneous complex. LARC binds to the hypersensitive 2 (HS2)-Maf recognition element (MARE) DNA in a sequence-specific manner and remodels nucleosomes. Heterogeneous nuclear ribonucleoprotein C1/C2, previously known as a general RNA binding protein, provides a sequence-specific DNA recognition element for LARC, and the LARC DNA-recognition sequence is essential for the enhancement of transcription by HS2. Independently of the initiation of transcription, LARC becomes associated with beta-like globin promoters.
Figures

Similar articles
-
Prevention of transcriptional silencing by a replicator-binding complex consisting of SWI/SNF, MeCP1, and hnRNP C1/C2.Mol Cell Biol. 2011 Aug;31(16):3472-84. doi: 10.1128/MCB.05587-11. Epub 2011 Jun 20. Mol Cell Biol. 2011. PMID: 21690294 Free PMC article.
-
Essential role of NF-E2 in remodeling of chromatin structure and transcriptional activation of the epsilon-globin gene in vivo by 5' hypersensitive site 2 of the beta-globin locus control region.Mol Cell Biol. 1996 Nov;16(11):6055-64. doi: 10.1128/MCB.16.11.6055. Mol Cell Biol. 1996. PMID: 8887635 Free PMC article.
-
Functional selectivity of recombinant mammalian SWI/SNF subunits.Genes Dev. 2000 Oct 1;14(19):2441-51. doi: 10.1101/gad.828000. Genes Dev. 2000. PMID: 11018012 Free PMC article.
-
Nucleosome organization and targeting of SWI/SNF chromatin-remodeling complexes: contributions of the DNA sequence.Biochem Cell Biol. 2007 Aug;85(4):419-25. doi: 10.1139/O07-070. Biochem Cell Biol. 2007. PMID: 17713577 Review.
-
ChIPs of the beta-globin locus: unraveling gene regulation within an active domain.Curr Opin Genet Dev. 2002 Apr;12(2):170-7. doi: 10.1016/s0959-437x(02)00283-6. Curr Opin Genet Dev. 2002. PMID: 11893490 Review.
Cited by
-
Towards a genome-wide reconstruction of cis-regulatory networks in the human genome.Semin Cell Dev Biol. 2009 Sep;20(7):842-8. doi: 10.1016/j.semcdb.2009.06.005. Epub 2009 Jun 26. Semin Cell Dev Biol. 2009. PMID: 19560550 Free PMC article. Review.
-
BRG1 requirement for long-range interaction of a locus control region with a downstream promoter.Proc Natl Acad Sci U S A. 2009 Feb 17;106(7):2259-64. doi: 10.1073/pnas.0806420106. Epub 2009 Jan 26. Proc Natl Acad Sci U S A. 2009. PMID: 19171905 Free PMC article.
-
Heterogeneous ribonucleoprotein C displays a repressor activity mediated by T-cell intracellular antigen-1-related/like protein to modulate Fas exon 6 splicing through a mechanism involving Hu antigen R.Nucleic Acids Res. 2010 Dec;38(22):8001-14. doi: 10.1093/nar/gkq698. Epub 2010 Aug 10. Nucleic Acids Res. 2010. PMID: 20699271 Free PMC article.
-
NPM and BRG1 Mediate Transcriptional Resistance to Retinoic Acid in Acute Promyelocytic Leukemia.Cell Rep. 2016 Mar 29;14(12):2938-49. doi: 10.1016/j.celrep.2016.02.074. Epub 2016 Mar 17. Cell Rep. 2016. PMID: 26997274 Free PMC article.
-
Mbd3/NURD complex regulates expression of 5-hydroxymethylcytosine marked genes in embryonic stem cells.Cell. 2011 Dec 23;147(7):1498-510. doi: 10.1016/j.cell.2011.11.054. Cell. 2011. PMID: 22196727 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
