

Without nerves, immunology remains incomplete -in vivo veritas
- PMID: 16162264
- PMCID: PMC1817817
- DOI: 10.1111/j.1365-2567.2005.02223.x
Without nerves, immunology remains incomplete -in vivo veritas
Abstract
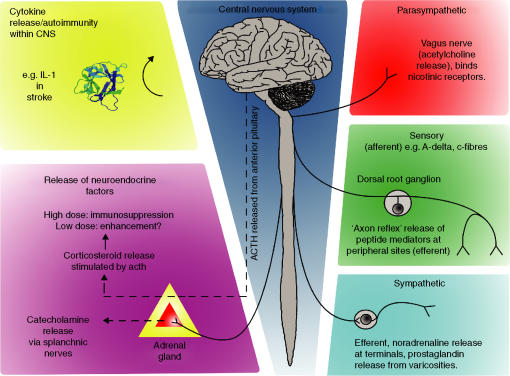
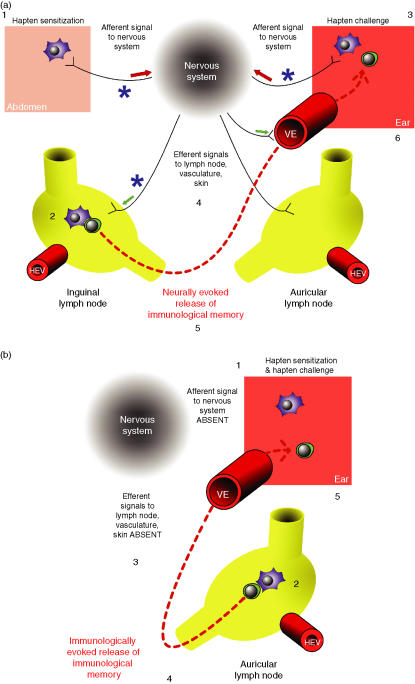
Interest in the interactions between nervous and immune systems involved in both pathological and homeostatic mechanisms of host defence has prompted studies of neuroendocrine immune modulation and cytokine involvement in neuropathologies. In this review we concentrate on a distinct area of homeostatic control of both normal and abnormal host defence activity involving the network of peripheral c-fibre nerve fibres. These nerve fibres have long been recognized by dermatologists and gastroenterologists as key players in abnormal inflammatory processes, such as dermatitis and eczema. However, the involvement of nerves can all too easily be regarded as that of isolated elements in a local phenomenon. On the contrary, it is becoming increasingly clear that neural monitoring of host defence activities takes place, and that involvement of central/spinal mechanisms are crucial in the co-ordination of the adaptive response to host challenge. We describe studies demonstrating neural control of host defence and use the specific examples of bone marrow haemopoiesis and contact sensitivity to highlight the role of direct nerve fibre connections in these activities. We propose a host monitoring system that requires interaction between specialized immune cells and nerve fibres distributed throughout the body and that gives rise to both neural and immune memories of prior challenge. While immunological mechanisms alone may be sufficient for local responsiveness to subsequent challenge, data are discussed that implicate the neural memory in co-ordination of host defence across the body, at distinct sites not served by the same nerve fibres, consistent with central nervous mediation.
Figures
Similar articles
-
Neural immunoregulation: emerging roles for nerves in immune homeostasis and disease.Immunol Today. 2000 Jun;21(6):281-9. doi: 10.1016/s0167-5699(00)01635-2. Immunol Today. 2000. PMID: 10825740 Review.
-
Central neural activation following contact sensitivity peripheral immune challenge: evidence of brain-immune regulation through C fibres.Immunology. 2015 Oct;146(2):206-16. doi: 10.1111/imm.12479. Epub 2015 Aug 24. Immunology. 2015. PMID: 25967648 Free PMC article.
-
The Regulation of Immunological Processes by Peripheral Neurons in Homeostasis and Disease.Trends Immunol. 2015 Oct;36(10):578-604. doi: 10.1016/j.it.2015.08.007. Trends Immunol. 2015. PMID: 26431937 Free PMC article. Review.
-
Neuroimmune communication in skin: far from peripheral.J Invest Dermatol. 2008 Feb;128(2):260-1. doi: 10.1038/sj.jid.5701171. J Invest Dermatol. 2008. PMID: 18195741 Review.
-
Integrative neuroimmunomodulation of gastrointestinal function during enteric parasitism.J Parasitol. 2001 Jun;87(3):483-504. doi: 10.1645/0022-3395(2001)087[0483:INOGFD]2.0.CO;2. J Parasitol. 2001. PMID: 11426710 Review.
Cited by
-
Peripheral nerve repair is associated with augmented cross-tissue inflammation following vascularized composite allotransplantation.Front Immunol. 2023 May 11;14:1151824. doi: 10.3389/fimmu.2023.1151824. eCollection 2023. Front Immunol. 2023. PMID: 37251389 Free PMC article.
-
Enteric ganglionitis in rhesus macaques infected with simian immunodeficiency virus.J Virol. 2007 Jun;81(12):6265-75. doi: 10.1128/JVI.02671-06. Epub 2007 Mar 28. J Virol. 2007. PMID: 17392357 Free PMC article.
-
The Interplay between Chronic Pain, Opioids, and the Immune System.Neuroscientist. 2022 Dec;28(6):613-627. doi: 10.1177/10738584211030493. Epub 2021 Jul 16. Neuroscientist. 2022. PMID: 34269117 Free PMC article. Review.
-
Neuroimmune Interaction in the Regulation of Peripheral Opioid-Mediated Analgesia in Inflammation.Front Immunol. 2016 Aug 2;7:293. doi: 10.3389/fimmu.2016.00293. eCollection 2016. Front Immunol. 2016. PMID: 27532001 Free PMC article. Review.
-
CGRP physiology, pharmacology, and therapeutic targets: migraine and beyond.Physiol Rev. 2023 Apr 1;103(2):1565-1644. doi: 10.1152/physrev.00059.2021. Epub 2022 Dec 1. Physiol Rev. 2023. PMID: 36454715 Free PMC article. Review.
References
-
- Lubahn CL, Schaller JA, Bellinger DL, Sweeney S, Lorton D. The importance of timing of adrenergic drug delivery in relation to the induction and onset of adjuvant-induced arthritis. Brain Behav Immun. 2004;18:563–71. - PubMed
-
- Harle P, Mobius D, Carr DJJ, Scholmerich J, Straub RH. An opposing time-dependent immune-modulating effect of the sympathetic nervous system conferred by altering the cytokine profile in the local lymph nodes and spleen of mice with type II collagen-induced arthritis. Arthritis Rheumatism. 2005;52:1305–13. - PubMed
-
- Sospedra M, Martin R. Immunology of multiple sclerosis. Annu Rev Immunol. 2005;23:683–747. - PubMed
-
- Gibson RM, Rothwell NJ, Le Feuvre RA. CNS injury: the role of the cytokine IL-1. Vet J. 2004;168:230–7. - PubMed
-
- Berczi I. Neuroimmune biology – an introduction. In: Berczi I, Gorczynski RM, editors. New Foundation of Biology. Oxford: Elsevier Science; 2001. pp. 1–46.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
