

Characterization of hortulanus endogenous murine leukemia virus, an endogenous provirus that encodes an infectious murine leukemia virus of a novel subgroup
- PMID: 15956577
- PMCID: PMC1143770
- DOI: 10.1128/JVI.79.13.8316-8329.2005
Characterization of hortulanus endogenous murine leukemia virus, an endogenous provirus that encodes an infectious murine leukemia virus of a novel subgroup
Abstract
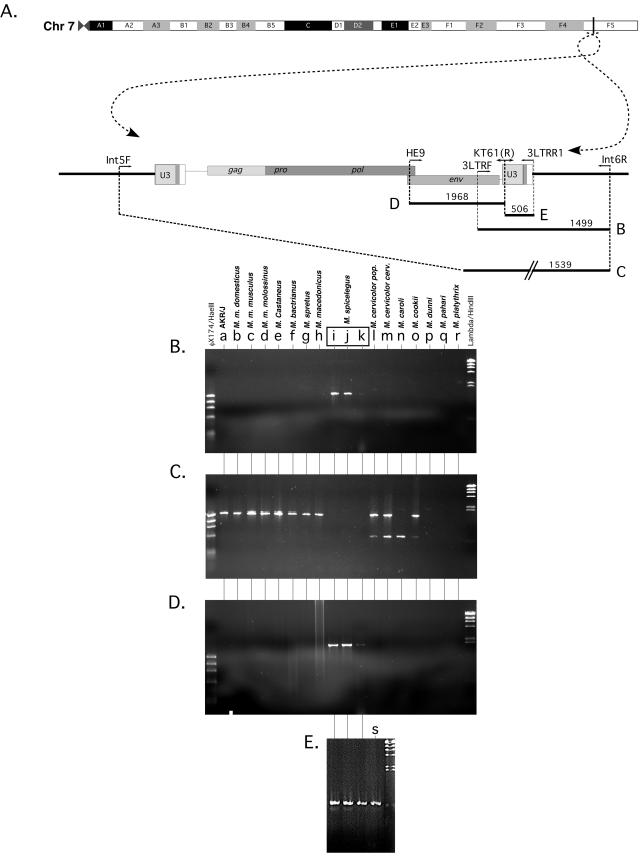
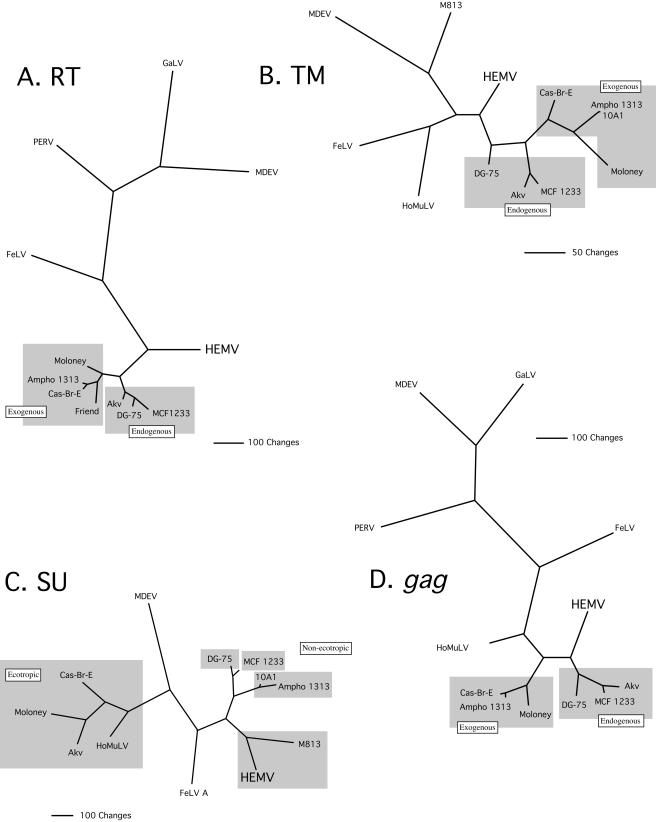
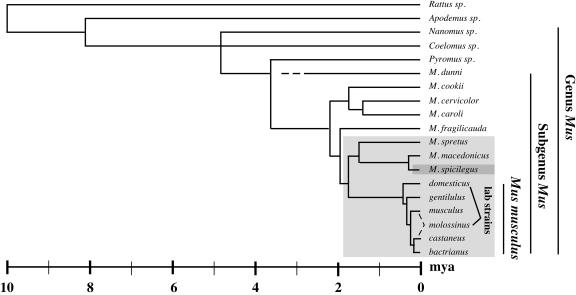
Simple retroviruses present a unique opportunity for examining the host-virus relationship. Following exogenous infection and integration into the germ line, copies of these viruses can become fixed within the genome. The resulting endogenous proviral "fossils" represent a record of past retroviral infections and forms. Previous work in our laboratory has been directed at dissecting the extensive nonecotropic murine leukemia virus content of the mouse genome. One such provirus, hortulanus endogenous murine leukemia virus (HEMV), found in a single copy in the genome of Mus spicilegus, was remarkable for characteristics that suggested that it was ancient and related to the hypothetical common ancestor of murine leukemia viruses (MLVs) and other gammaretroviral species. In the present study, we have analyzed its functional properties. Transfection of a molecular clone of the HEMV provirus into mouse-derived cell lines revealed that it is replication competent. Furthermore, host range and interference studies revealed a strictly ecotropic host range and the use of a receptor distinct from those used by other classical MLVs. The identity of nucleotide sequence of the long terminal repeats (LTRs) further suggested that HEMV is a relatively recent insertion into the M. spicilegus genome at the distal end of chromosome 7. Although unique to M. spicilegus, its presence in a homozygous state in three individuals obtained from different regions implies that it has been present long enough to become fixed in this species. Exhaustive phylogenetic analysis of all regions of the HEMV genome supported the previously assigned ancestral position of HEMV relative to other MLV-related viruses. Thus, HEMV is a relatively recent introduction into the Mus germ line but is representative of a relatively ancestral MLV group.
Figures

Similar articles
-
Structures of endogenous nonecotropic murine leukemia virus (MLV) long terminal repeats in wild mice: implication for evolution of MLVs.J Virol. 1999 May;73(5):4327-40. doi: 10.1128/JVI.73.5.4327-4340.1999. J Virol. 1999. PMID: 10196331 Free PMC article.
-
Rmcf2, a xenotropic provirus in the Asian mouse species Mus castaneus, blocks infection by polytropic mouse gammaretroviruses.J Virol. 2005 Aug;79(15):9677-84. doi: 10.1128/JVI.79.15.9677-9684.2005. J Virol. 2005. PMID: 16014929 Free PMC article.
-
Antigenic subclasses of polytropic murine leukemia virus (MLV) isolates reflect three distinct groups of endogenous polytropic MLV-related sequences in NFS/N mice.J Virol. 2003 Oct;77(19):10327-38. doi: 10.1128/jvi.77.19.10327-10338.2003. J Virol. 2003. PMID: 12970417 Free PMC article.
-
Origins of the endogenous and infectious laboratory mouse gammaretroviruses.Viruses. 2014 Dec 26;7(1):1-26. doi: 10.3390/v7010001. Viruses. 2014. PMID: 25549291 Free PMC article. Review.
-
Endogenous retroviruses and the human germline.Curr Opin Genet Dev. 2000 Dec;10(6):651-5. doi: 10.1016/s0959-437x(00)00138-6. Curr Opin Genet Dev. 2000. PMID: 11088016 Review.
Cited by
-
Mus spicilegus endogenous retrovirus HEMV uses murine sodium-dependent myo-inositol transporter 1 as a receptor.J Virol. 2012 Jun;86(11):6341-4. doi: 10.1128/JVI.00083-12. Epub 2012 Mar 28. J Virol. 2012. PMID: 22457525 Free PMC article.
-
Murine gammaretrovirus group G3 was not found in Swedish patients with myalgic encephalomyelitis/chronic fatigue syndrome and fibromyalgia.PLoS One. 2011;6(10):e24602. doi: 10.1371/journal.pone.0024602. Epub 2011 Oct 12. PLoS One. 2011. PMID: 22022360 Free PMC article.
-
The GLN family of murine endogenous retroviruses contains an element competent for infectious viral particle formation.J Virol. 2008 May;82(9):4413-9. doi: 10.1128/JVI.02141-07. Epub 2008 Feb 20. J Virol. 2008. PMID: 18287236 Free PMC article.
-
New simian beta retroviruses from rhesus monkeys (Macaca mulatta) and langurs (Semnopithecus entellus) from Rajasthan, India.Virus Genes. 2006 Aug;33(1):107-16. doi: 10.1007/s11262-005-0032-x. Virus Genes. 2006. PMID: 16791425
-
Molecular and phylogenetic analyses of a new amphotropic murine leukemia virus (MuLV-1313).Virol J. 2006 Dec 5;3:101. doi: 10.1186/1743-422X-3-101. Virol J. 2006. PMID: 17147829 Free PMC article.
References
-
- Albritton, L. M., L. Tseng, D. Scadden, and J. M. Cunningham. 1989. A putative murine ecotropic retrovirus receptor gene encodes a multiple membrane-spanning protein and confers susceptibility to virus infection. Cell 57:659-666. - PubMed
-
- Auffray, J., A. Orth, J. Catalan, J. Gonzalez, E. Desmarais, and F. Bonhomme. 2003. Phylogenetic position and description of a new species of subgenus Mus (Rodentia, Mammalia) from Thailand. Zoologica Scripta 32:119-127.
-
- Bonhomme, F., and J. Guenet. 1995. The Laboratory mouse and its wild relatives, 3rd ed. Oxford University Press, New York, N.Y.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
