

Control of translation by the 5'- and 3'-terminal regions of the dengue virus genome
- PMID: 15956576
- PMCID: PMC1143759
- DOI: 10.1128/JVI.79.13.8303-8315.2005
Control of translation by the 5'- and 3'-terminal regions of the dengue virus genome
Abstract
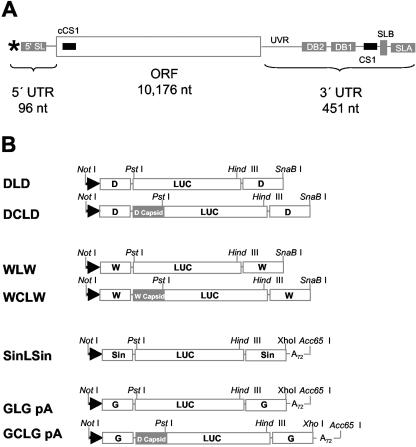
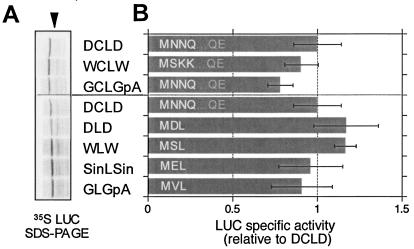
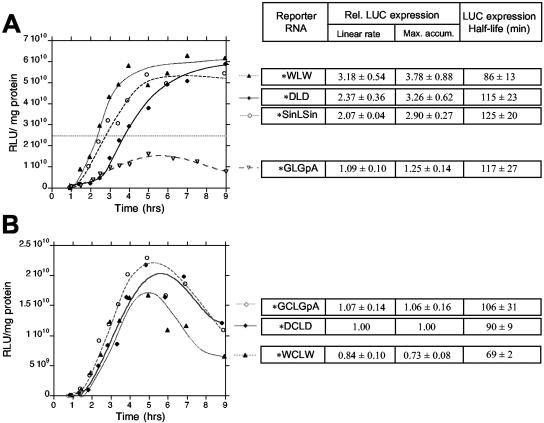
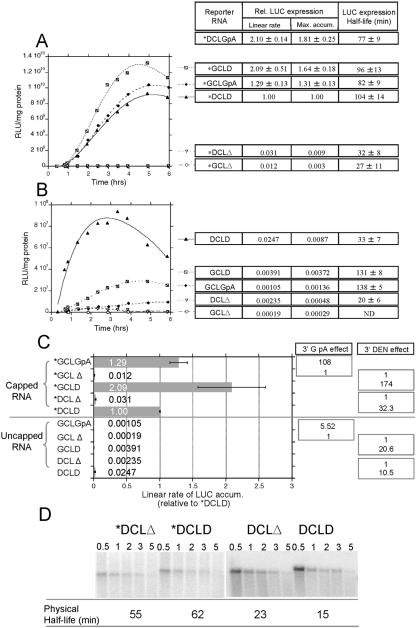
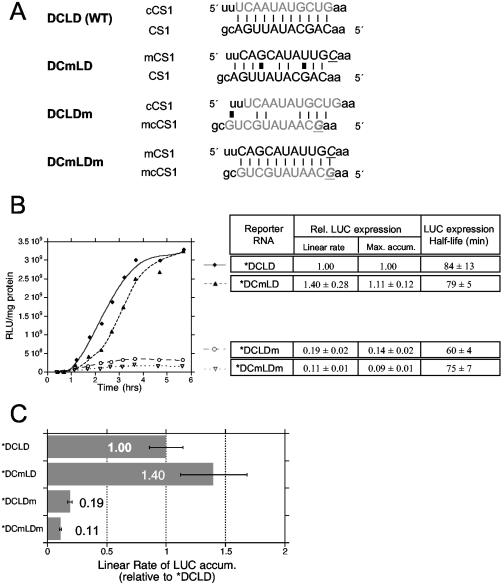
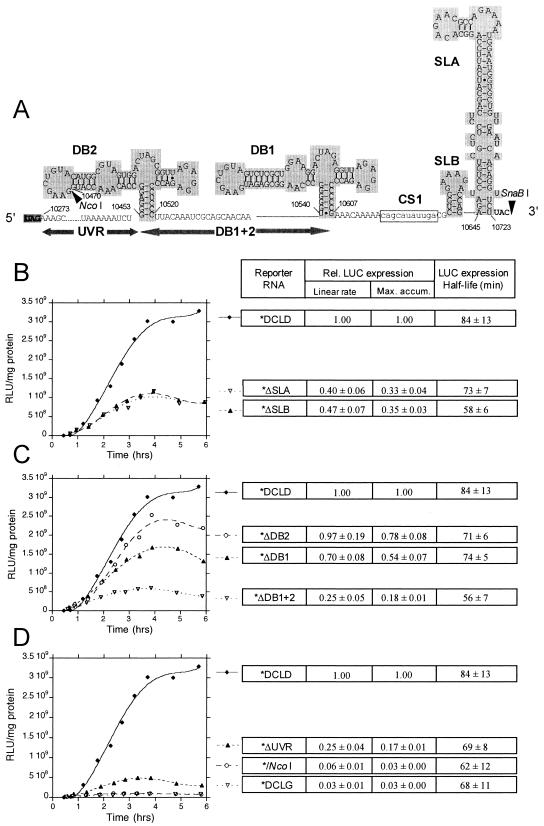
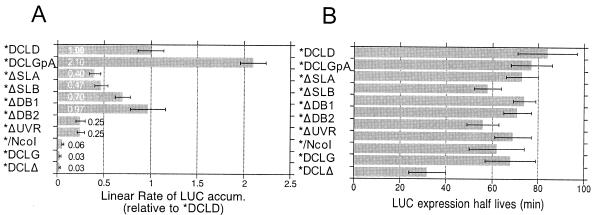
The genomic RNAs of flaviviruses such as dengue virus (DEN) have a 5' m7GpppN cap like those of cellular mRNAs but lack a 3' poly(A) tail. We have studied the contributions to translational expression of 5'- and 3'-terminal regions of the DEN serotype 2 genome by using luciferase reporter mRNAs transfected into Vero cells. DCLD RNA contained the entire DEN 5' and 3' untranslated regions (UTRs), as well as the first 36 codons of the capsid coding region fused to the luciferase reporter gene. Capped DCLD RNA was as efficiently translated in Vero cells as capped GLGpA RNA, a reporter with UTRs from the highly expressed alpha-globin mRNA and a 72-residue poly(A) tail. Analogous reporter RNAs with regulatory sequences from West Nile and Sindbis viruses were also strongly expressed. Although capped DCLD RNA was expressed much more efficiently than its uncapped form, uncapped DCLD RNA was translated 6 to 12 times more efficiently than uncapped RNAs with UTRs from globin mRNA. The 5' cap and DEN 3' UTR were the main sources of the translational efficiency of DCLD RNA, and they acted synergistically in enhancing translation. The DEN 3' UTR increased mRNA stability, although this effect was considerably weaker than the enhancement of translational efficiency. The DEN 3' UTR thus has translational regulatory properties similar to those of a poly(A) tail. Its translation-enhancing effect was observed for RNAs with globin or DEN 5' sequences, indicating no codependency between viral 5' and 3' sequences. Deletion studies showed that translational enhancement provided by the DEN 3' UTR is attributable to the cumulative contributions of several conserved elements, as well as a nonconserved domain adjacent to the stop codon. One of the conserved elements was the conserved sequence (CS) CS1 that is complementary to cCS1 present in the 5' end of the DEN polyprotein open reading frame. Complementarity between CS1 and cCS1 was not required for efficient translation.
Figures
Similar articles
-
Dengue virus utilizes a novel strategy for translation initiation when cap-dependent translation is inhibited.J Virol. 2006 Mar;80(6):2976-86. doi: 10.1128/JVI.80.6.2976-2986.2006. J Virol. 2006. PMID: 16501107 Free PMC article.
-
Enhancement of dengue virus translation: role of the 3' untranslated region and the terminal 3' stem-loop domain.Virology. 2004 Nov 10;329(1):119-33. doi: 10.1016/j.virol.2004.08.004. Virology. 2004. PMID: 15476880
-
Untranslated regions of diverse plant viral RNAs vary greatly in translation enhancement efficiency.BMC Biotechnol. 2012 May 6;12:22. doi: 10.1186/1472-6750-12-22. BMC Biotechnol. 2012. PMID: 22559081 Free PMC article.
-
[Structure and function of the non-coding regions of hepatitis C viral RNA].Postepy Biochem. 2006;52(1):62-71. Postepy Biochem. 2006. PMID: 16869303 Review. Polish.
-
End-to-end communication in the modulation of translation by mammalian RNA viruses.Virus Res. 2006 Jul;119(1):43-51. doi: 10.1016/j.virusres.2005.10.012. Epub 2005 Nov 22. Virus Res. 2006. PMID: 16307817 Free PMC article. Review.
Cited by
-
Evidence for inter- and intra-genotypic variations in dengue serotype 4 viruses representing predominant and non-predominant genotypes co-circulating in Thailand from 1977 to 2001.Virus Genes. 2010 Aug;41(1):5-13. doi: 10.1007/s11262-010-0473-8. Epub 2010 Mar 25. Virus Genes. 2010. PMID: 20336481
-
Characterization of cis-Acting RNA Elements of Zika Virus by Using a Self-Splicing Ribozyme-Dependent Infectious Clone.J Virol. 2017 Oct 13;91(21):e00484-17. doi: 10.1128/JVI.00484-17. Print 2017 Nov 1. J Virol. 2017. PMID: 28814522 Free PMC article.
-
Animal virus schemes for translation dominance.Curr Opin Virol. 2011 Nov;1(5):363-72. doi: 10.1016/j.coviro.2011.10.009. Curr Opin Virol. 2011. PMID: 22319551 Free PMC article. Review.
-
Structures required for poly(A) tail-independent translation overlap with, but are distinct from, cap-independent translation and RNA replication signals at the 3' end of Tobacco necrosis virus RNA.Virology. 2007 Feb 20;358(2):448-58. doi: 10.1016/j.virol.2006.08.054. Epub 2006 Oct 4. Virology. 2007. PMID: 17023016 Free PMC article.
-
Viral strategies to subvert the mammalian translation machinery.Prog Mol Biol Transl Sci. 2009;90:313-67. doi: 10.1016/S1877-1173(09)90009-6. Epub 2009 Oct 27. Prog Mol Biol Transl Sci. 2009. PMID: 20374746 Free PMC article. Review.
References
-
- Brinton, M. A., and J. H. Dispoto. 1988. Sequence and secondary structure analysis of the 5′-terminal region of flavivirus genome RNA. Virology 162:290-299. - PubMed
-
- Brinton, M. A., A. V. Fernandez, and J. H. Dispoto. 1986. The 3′-nucleotides of flavivirus genomic RNA form a conserved secondary structure. Virology 153:113-121. - PubMed
-
- Burke, D. S., and T. P. Monath. 2001. Flaviviruses, p. 1043-1125. In D. M. Knipe and P. M. Howley (ed.), Fields virology, 4th ed. Lippincott Williams & Wilkins, Philadelphia, Pa.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
