

Normal microRNA maturation and germ-line stem cell maintenance requires Loquacious, a double-stranded RNA-binding domain protein
- PMID: 15918770
- PMCID: PMC1141267
- DOI: 10.1371/journal.pbio.0030236
Normal microRNA maturation and germ-line stem cell maintenance requires Loquacious, a double-stranded RNA-binding domain protein
Abstract
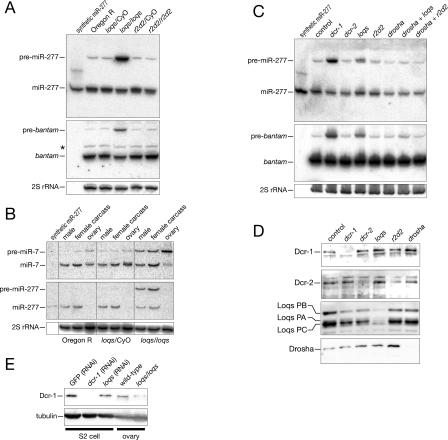
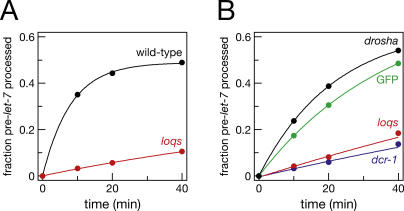
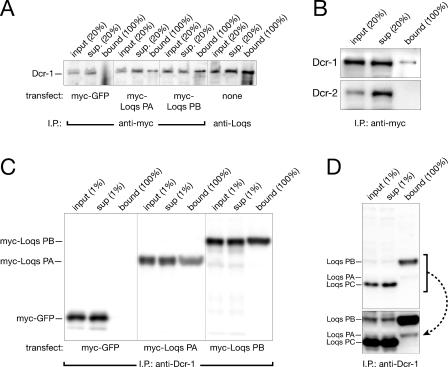
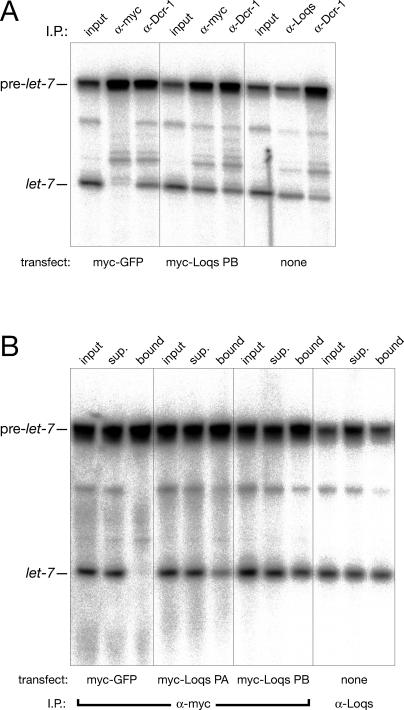
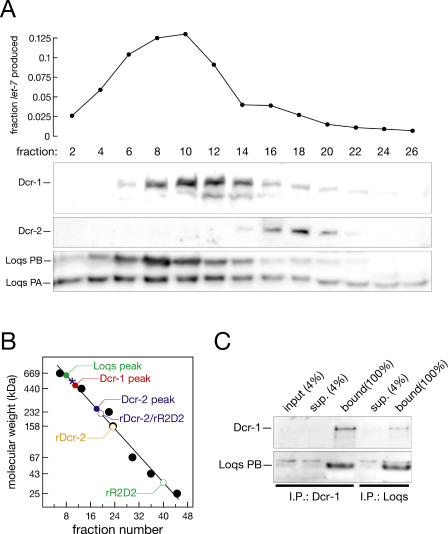
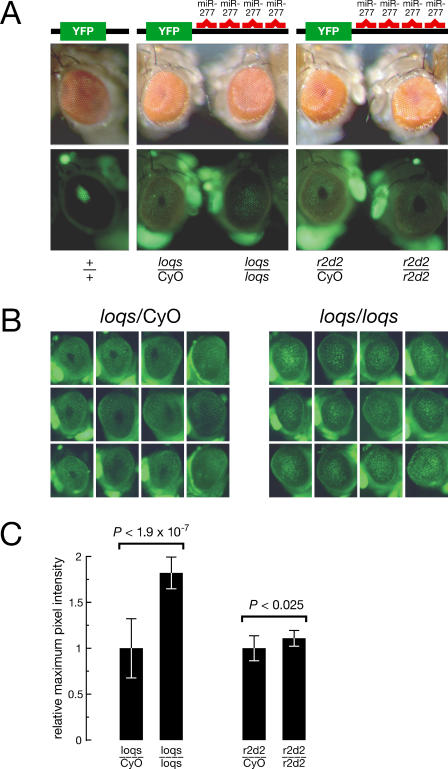
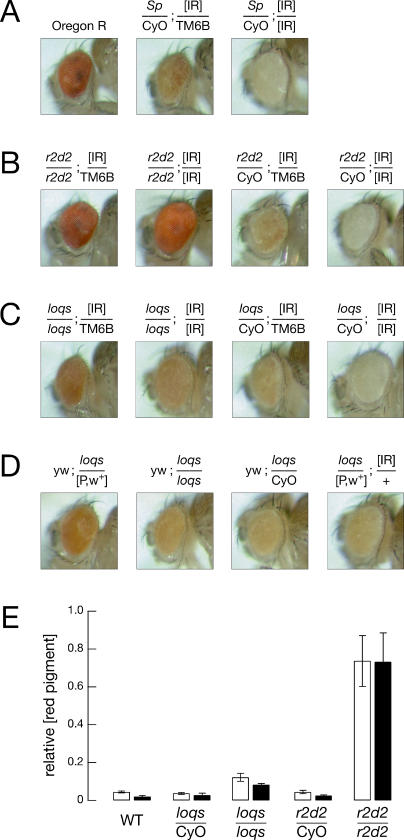
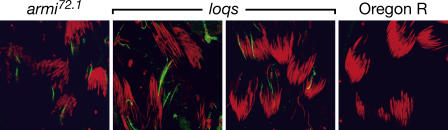
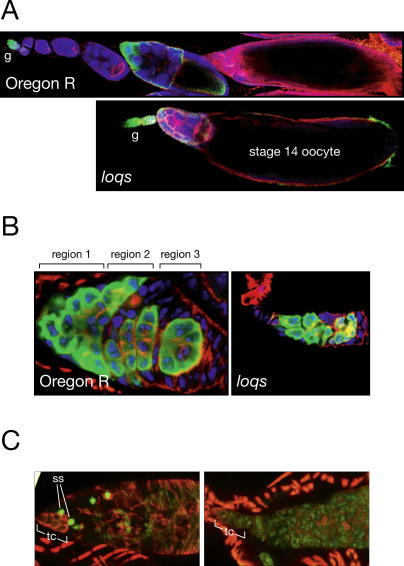
microRNAs (miRNAs) are single-stranded, 21- to 23-nucleotide cellular RNAs that control the expression of cognate target genes. Primary miRNA (pri-miRNA) transcripts are transformed to mature miRNA by the successive actions of two RNase III endonucleases. Drosha converts pri-miRNA transcripts to precursor miRNA (pre-miRNA); Dicer, in turn, converts pre-miRNA to mature miRNA. Here, we show that normal processing of Drosophila pre-miRNAs by Dicer-1 requires the double-stranded RNA-binding domain (dsRBD) protein Loquacious (Loqs), a homolog of human TRBP, a protein first identified as binding the HIV trans-activator RNA (TAR). Efficient miRNA-directed silencing of a reporter transgene, complete repression of white by a dsRNA trigger, and silencing of the endogenous Stellate locus by Suppressor of Stellate, all require Loqs. In loqs(f00791) mutant ovaries, germ-line stem cells are not appropriately maintained. Loqs associates with Dcr-1, the Drosophila RNase III enzyme that processes pre-miRNA into mature miRNA. Thus, every known Drosophila RNase-III endonuclease is paired with a dsRBD protein that facilitates its function in small RNA biogenesis.
Figures

Similar articles
-
Processing of pre-microRNAs by the Dicer-1-Loquacious complex in Drosophila cells.PLoS Biol. 2005 Jul;3(7):e235. doi: 10.1371/journal.pbio.0030235. Epub 2005 May 24. PLoS Biol. 2005. PMID: 15918769 Free PMC article.
-
Functional anatomy of the Drosophila microRNA-generating enzyme.J Biol Chem. 2007 Sep 28;282(39):28373-28378. doi: 10.1074/jbc.M705208200. Epub 2007 Jul 31. J Biol Chem. 2007. PMID: 17666393
-
Dicer-1, but not Loquacious, is critical for assembly of miRNA-induced silencing complexes.RNA. 2007 Dec;13(12):2324-9. doi: 10.1261/rna.723707. Epub 2007 Oct 10. RNA. 2007. PMID: 17928574 Free PMC article.
-
MicroRNAs: Loquacious speaks out.Curr Biol. 2005 Aug 9;15(15):R603-5. doi: 10.1016/j.cub.2005.07.044. Curr Biol. 2005. PMID: 16085484 Review.
-
The double-stranded microRNA precursor.Postepy Biochem. 2024 May 23;70(1):57-61. doi: 10.18388/pb.2021_522. Print 2024 May 23. Postepy Biochem. 2024. PMID: 39016229 Review.
Cited by
-
Differential roles of human Dicer-binding proteins TRBP and PACT in small RNA processing.Nucleic Acids Res. 2013 Jul;41(13):6568-76. doi: 10.1093/nar/gkt361. Epub 2013 May 9. Nucleic Acids Res. 2013. PMID: 23661684 Free PMC article.
-
Mei-P26 regulates the maintenance of ovarian germline stem cells by promoting BMP signaling.Development. 2012 May;139(9):1547-56. doi: 10.1242/dev.077412. Epub 2012 Mar 21. Development. 2012. PMID: 22438571 Free PMC article.
-
Myogenic factors that regulate expression of muscle-specific microRNAs.Proc Natl Acad Sci U S A. 2006 Jun 6;103(23):8721-6. doi: 10.1073/pnas.0602831103. Epub 2006 May 26. Proc Natl Acad Sci U S A. 2006. PMID: 16731620 Free PMC article.
-
Telomeric trans-silencing: an epigenetic repression combining RNA silencing and heterochromatin formation.PLoS Genet. 2007 Sep;3(9):1633-43. doi: 10.1371/journal.pgen.0030158. Epub 2007 Jul 31. PLoS Genet. 2007. PMID: 17941712 Free PMC article.
-
The Arabidopsis thaliana double-stranded RNA binding protein DRB1 directs guide strand selection from microRNA duplexes.RNA. 2009 Dec;15(12):2219-35. doi: 10.1261/rna.1646909. Epub 2009 Oct 27. RNA. 2009. PMID: 19861421 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
