

Review
doi: 10.1146/annurev.immunol.23.021704.115653.
How neutrophils kill microbes
Affiliations
- PMID: 15771570
- PMCID: PMC2092448
- DOI: 10.1146/annurev.immunol.23.021704.115653
Item in Clipboard
Review
How neutrophils kill microbes
Annu Rev Immunol.
2005.
Abstract
Neutrophils provide the first line of defense of the innate immune system by phagocytosing, killing, and digesting bacteria and fungi. Killing was previously believed to be accomplished by oxygen free radicals and other reactive oxygen species generated by the NADPH oxidase, and by oxidized halides produced by myeloperoxidase. We now know this is incorrect. The oxidase pumps electrons into the phagocytic vacuole, thereby inducing a charge across the membrane that must be compensated. The movement of compensating ions produces conditions in the vacuole conducive to microbial killing and digestion by enzymes released into the vacuole from the cytoplasmic granules.
Figures
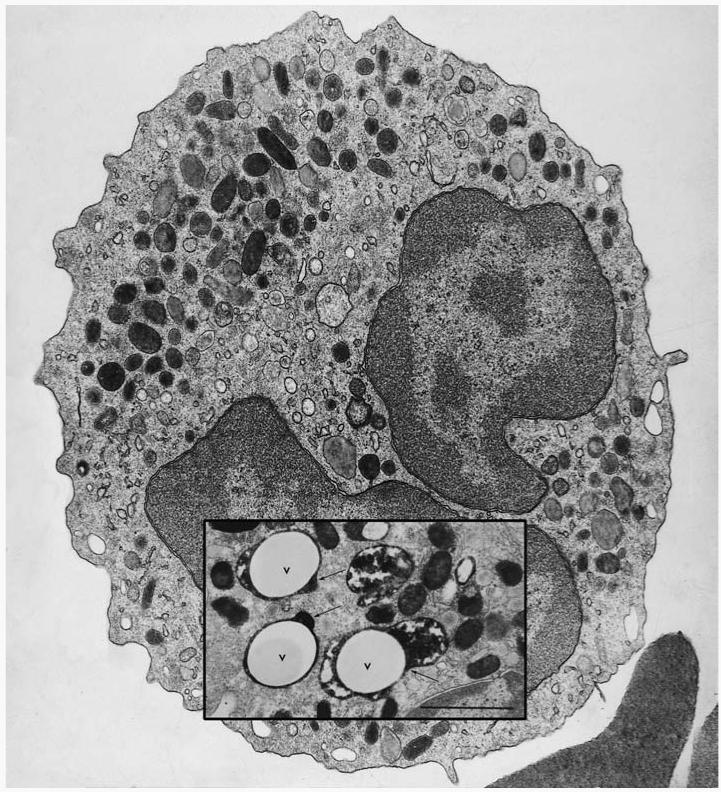
Transmission electron micrograph of a human neutrophil. Inset is an image taken from a neutrophil 20 s after the phagocytosis of latex particles opsonized with IgG (V, vacuole). The section was stained for myeloperoxidase (MPO) to reveal the electron-dense product in the azurophil granules, some of which can be seen degranulating into the phagocytic vacuole (arrows). Bar = 1 μm. (Figure from .)

Schematic representation of the NADPH oxidase. Flavocytochrome b558 is a heterodimer of gp91phox, which contains the haem- and flavin-binding sites, and p22phox. Electron transport is activated by phosphorylation and translocation to the vacuolar membrane of p47phox and p67phox. p21rac, in the GTP-bound form, is also required (12).
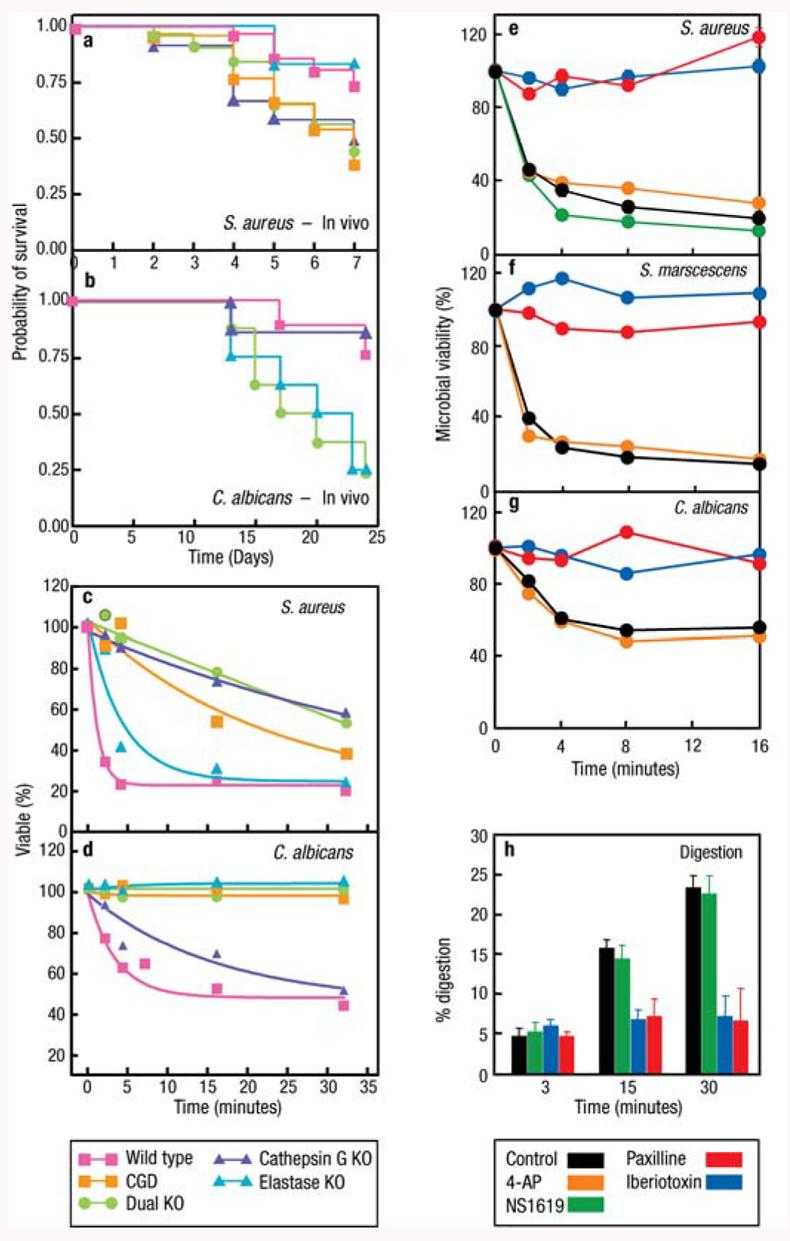
The neutral proteases elastase and cathepsin G as well as K+ flux are required for microbial killing and digestion by neutrophils. Cathepsin G, neutrophil elastase (NE), and p47phox (CGD) knockout mice are susceptible to S. aureus (a) and C. albicans (b) in vivo, and their neutrophils kill these organisms poorly in the test tube (c) and (d) (adapted from 6). Inhibition of the BKCa K+ channel with specific inhibitors paxilline (PAX) and iberiotoxin (IBTX) prevents killing of S. aureus (e), S. marscescens (f ), and C. albicans (g) by neutrophils, whereas the opener NS1619 and nonspecific inhibitor 4-aminopyridine were without effect. The BKCa K+ channel blockers also inhibited digestion of radiolabeled, killed S. aureus (h) (adapted from 74). Neither the loss of the proteases nor blockage of the BKCa channel affected phagocytosis, oxidase activity, or iodination.
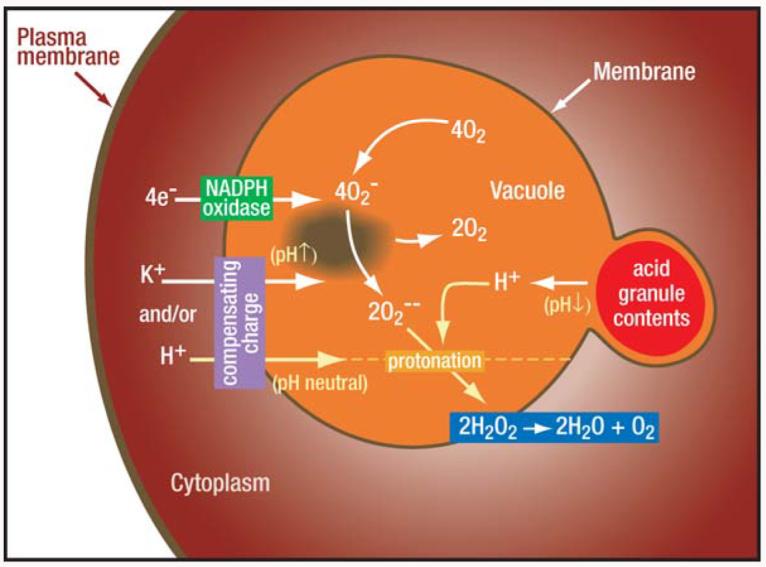
Activity of the NADPH oxidase depolarizes the membrane. The nature of the compensating charge governs the changes in vacuolar pH and tonicity. Electrons are transported across the vacuolar membrane to form , which dismutates to . and become protonated to form HO2 and H2O2, thereby consuming protons and elevating the pH in the vacuole despite the entry of acidic granule contents. This process can only occur if part of the charge is compensated by ions other than protons, which in part occurs through the passage of K+ ions (6, 74).
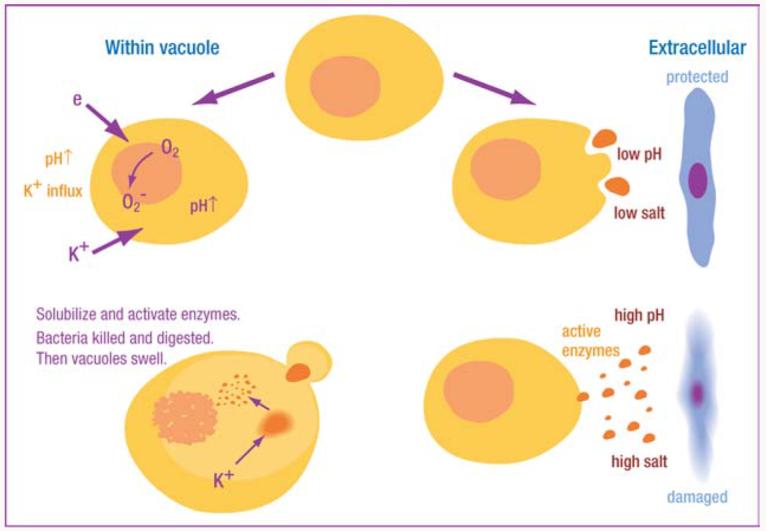
Schematic representation of interaction between NADPH oxidase and granule proteases. Electron transport through flavocytochrome b558 consumes protons in the vacuole, elevating pH to a level optimal for neutral proteases, which are also activated by K+ driven into the vacuole to compensate the charge across the membrane. The hypertonic K+ solubilizes the cationic granule proteases and peptides by displacing them from the anionic sulphated proteoglycan granule matrix. The requirement for an alkaline, hypertonic environment restricts the toxicity of these proteins to the vacuolar compartment, thereby limiting damage to normal tissues.
Similar articles
-
How superoxide production by neutrophil leukocytes kills microbes.Novartis Found Symp. 2006;279:92-8; discussion 98-100, 216-9. Novartis Found Symp. 2006. PMID: 17278388
-
The NADPH Oxidase and Microbial Killing by Neutrophils, With a Particular Emphasis on the Proposed Antimicrobial Role of Myeloperoxidase within the Phagocytic Vacuole.Microbiol Spectr. 2016 Aug;4(4). doi: 10.1128/microbiolspec.MCHD-0018-2015. Microbiol Spectr. 2016. PMID: 27726789 Review.
-
Reassessment of the microbicidal activity of reactive oxygen species and hypochlorous acid with reference to the phagocytic vacuole of the neutrophil granulocyte.J Med Microbiol. 2003 Aug;52(Pt 8):643-651. doi: 10.1099/jmm.0.05181-0. J Med Microbiol. 2003. PMID: 12867557 Free PMC article.
-
Oxidative innate immune defenses by Nox/Duox family NADPH oxidases.Contrib Microbiol. 2008;15:164-187. doi: 10.1159/000136357. Contrib Microbiol. 2008. PMID: 18511861 Free PMC article. Review.
-
Pseudomonas aeruginosa Effector ExoS Inhibits ROS Production in Human Neutrophils.Cell Host Microbe. 2017 May 10;21(5):611-618.e5. doi: 10.1016/j.chom.2017.04.001. Cell Host Microbe. 2017. PMID: 28494242 Free PMC article.
Cited by
-
Cell Population Data (CPD) for Early Recognition of Sepsis and Septic Shock in Children: A Pilot Study.Front Pediatr. 2021 Mar 8;9:642377. doi: 10.3389/fped.2021.642377. eCollection 2021. Front Pediatr. 2021. PMID: 33777867 Free PMC article.
-
The genetic theory of infectious diseases: a brief history and selected illustrations.Annu Rev Genomics Hum Genet. 2013;14:215-43. doi: 10.1146/annurev-genom-091212-153448. Epub 2013 May 29. Annu Rev Genomics Hum Genet. 2013. PMID: 23724903 Free PMC article. Review.
-
CpsY influences Streptococcus iniae cell wall adaptations important for neutrophil intracellular survival.Infect Immun. 2012 May;80(5):1707-15. doi: 10.1128/IAI.00027-12. Epub 2012 Feb 21. Infect Immun. 2012. PMID: 22354020 Free PMC article.
-
Urban Particulate Matter Triggers Meibomian Gland Dysfunction.Invest Ophthalmol Vis Sci. 2024 Feb 1;65(2):8. doi: 10.1167/iovs.65.2.8. Invest Ophthalmol Vis Sci. 2024. PMID: 38315493 Free PMC article.
-
Interactions between polymorphonuclear leukocytes and Pseudomonas aeruginosa biofilms on silicone implants in vivo.Infect Immun. 2012 Aug;80(8):2601-7. doi: 10.1128/IAI.06215-11. Epub 2012 May 14. Infect Immun. 2012. PMID: 22585963 Free PMC article.
References
-
- Iyer GYN, Islam DMF, Quastel JH. Biochemical aspects of phagocytosis. Nature. 1961;192:535–41.
-
- Klebanoff SJ, White LR. Iodination defect in the leukocytes of a patient with chronic granulomatous disease of childhood. N. Engl. J. Med. 1969;280:460–66. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
