

An early step in wobble uridine tRNA modification requires the Elongator complex
- PMID: 15769872
- PMCID: PMC1370732
- DOI: 10.1261/rna.7247705
An early step in wobble uridine tRNA modification requires the Elongator complex
Abstract
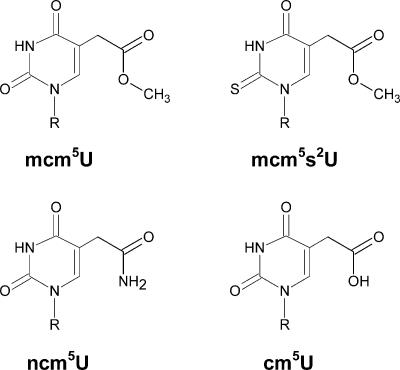
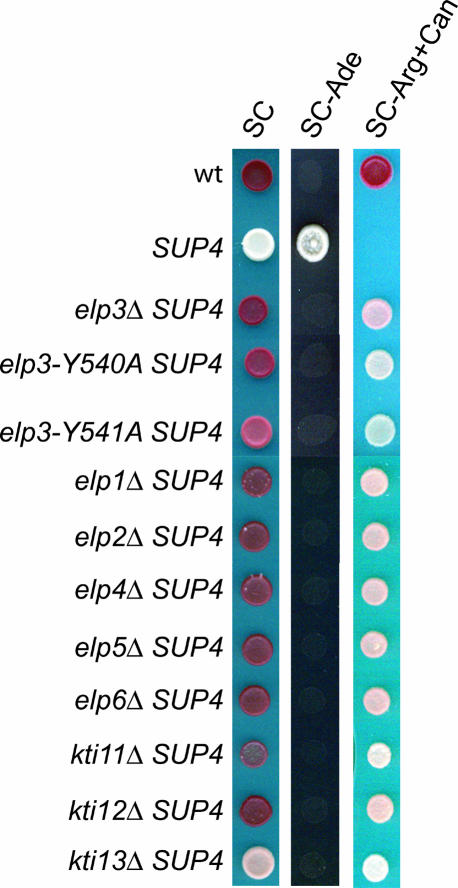
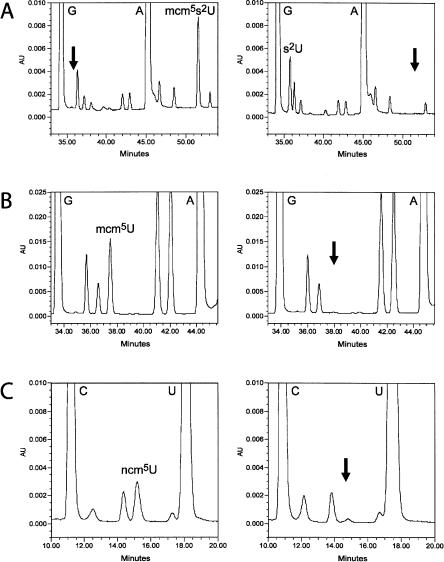
Elongator has been reported to be a histone acetyltransferase complex involved in elongation of RNA polymerase II transcription. In Saccharomyces cerevisiae, mutations in any of the six Elongator protein subunit (ELP1-ELP6) genes or the three killer toxin insensitivity (KTI11-KTI13) genes cause similar pleiotropic phenotypes. By analyzing modified nucleosides in individual tRNA species, we show that the ELP1-ELP6 and KTI11-KTI13 genes are all required for an early step in synthesis of 5-methoxycarbonylmethyl (mcm5) and 5-carbamoylmethyl (ncm5) groups present on uridines at the wobble position in tRNA. Transfer RNA immunoprecipitation experiments showed that the Elp1 and Elp3 proteins specifically coprecipitate a tRNA susceptible to formation of an mcm5 side chain, indicating a direct role of Elongator in tRNA modification. The presence of mcm5U, ncm5U, or derivatives thereof at the wobble position is required for accurate and efficient translation, suggesting that the phenotypes of elp1-elp6 and kti11-kti13 mutants could be caused by a translational defect. Accordingly, a deletion of any ELP1-ELP6 or KTI11-KTI13 gene prevents an ochre suppressor tRNA that normally contains mcm5U from reading ochre stop codons.
Figures

Similar articles
-
Yeast alpha-tubulin suppressor Ats1/Kti13 relates to the Elongator complex and interacts with Elongator partner protein Kti11.Mol Microbiol. 2008 Jul;69(1):175-87. doi: 10.1111/j.1365-2958.2008.06273.x. Epub 2008 May 5. Mol Microbiol. 2008. PMID: 18466297
-
Elevated levels of two tRNA species bypass the requirement for elongator complex in transcription and exocytosis.Mol Cell. 2006 Oct 6;24(1):139-48. doi: 10.1016/j.molcel.2006.07.031. Mol Cell. 2006. PMID: 17018299
-
Structure of the Elongator cofactor complex Kti11/Kti13 provides insight into the role of Kti13 in Elongator-dependent tRNA modification.FEBS J. 2015 Mar;282(5):819-33. doi: 10.1111/febs.13199. Epub 2015 Feb 4. FEBS J. 2015. PMID: 25604895
-
Elongator, a conserved complex required for wobble uridine modifications in eukaryotes.RNA Biol. 2014;11(12):1519-28. doi: 10.4161/15476286.2014.992276. RNA Biol. 2014. PMID: 25607684 Free PMC article. Review.
-
Elongator-a tRNA modifying complex that promotes efficient translational decoding.Biochim Biophys Acta Gene Regul Mech. 2018 Apr;1861(4):401-408. doi: 10.1016/j.bbagrm.2017.11.006. Epub 2017 Nov 21. Biochim Biophys Acta Gene Regul Mech. 2018. PMID: 29170010 Review.
Cited by
-
Variation of tRNA modifications with and without intron dependency.Front Genet. 2024 Sep 4;15:1460902. doi: 10.3389/fgene.2024.1460902. eCollection 2024. Front Genet. 2024. PMID: 39296543 Free PMC article. Review.
-
RNA-modifying enzyme Alkbh8 is involved in mouse embryonic development.iScience. 2024 Aug 22;27(9):110777. doi: 10.1016/j.isci.2024.110777. eCollection 2024 Sep 20. iScience. 2024. PMID: 39280612 Free PMC article.
-
ELP1, the Gene Mutated in Familial Dysautonomia, Is Required for Normal Enteric Nervous System Development and Maintenance and for Gut Epithelium Homeostasis.J Neurosci. 2024 Sep 11;44(37):e2253232024. doi: 10.1523/JNEUROSCI.2253-23.2024. J Neurosci. 2024. PMID: 39138000
-
Mechanisms and Delivery of tRNA Therapeutics.Chem Rev. 2024 Jun 26;124(12):7976-8008. doi: 10.1021/acs.chemrev.4c00142. Epub 2024 May 27. Chem Rev. 2024. PMID: 38801719 Review.
-
Cryo-EM structures of the human Elongator complex at work.Nat Commun. 2024 May 15;15(1):4094. doi: 10.1038/s41467-024-48251-y. Nat Commun. 2024. PMID: 38750017 Free PMC article.
References
-
- Agarwalla, S., Kealey, J.T., Santi, D.V., and Stroud, R.M. 2002. Characterization of the 23 S ribosomal RNA m5U1939 methyltransferase from Escherichia coli. J. Biol. Chem. 277: 8835–8840. - PubMed
-
- Agris, P.F. 1991. Wobble position modified nucleosides evolved to select transfer RNA codon recognition: A modified-wobble hypothesis. Biochimie 73: 1345–1349. - PubMed
-
- Amstutz, H., Munz, P., Heyer, W.D., Leupoid, U., and Kohli, J. 1985. Concerted evolution of tRNA genes: Intergenic conversion among three unlinked serine tRNA genes in S. pombe. Cell 40: 879–886. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases