

RGG-boxes of the EWS oncoprotein repress a range of transcriptional activation domains
- PMID: 15743974
- PMCID: PMC552958
- DOI: 10.1093/nar/gki270
RGG-boxes of the EWS oncoprotein repress a range of transcriptional activation domains
Abstract
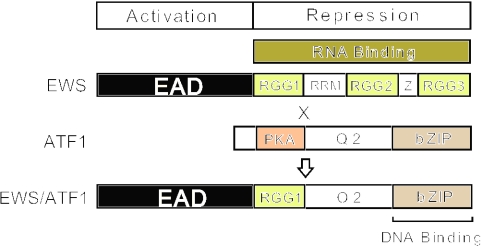
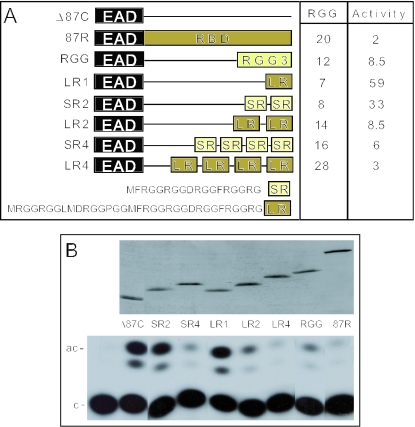
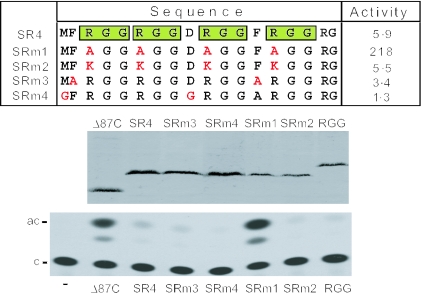
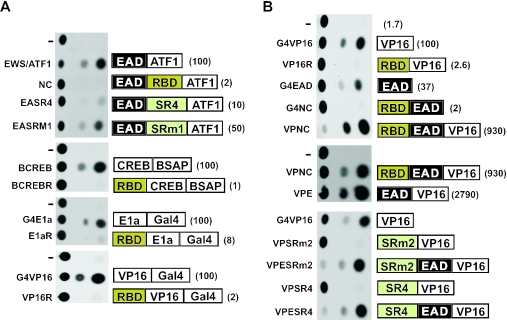
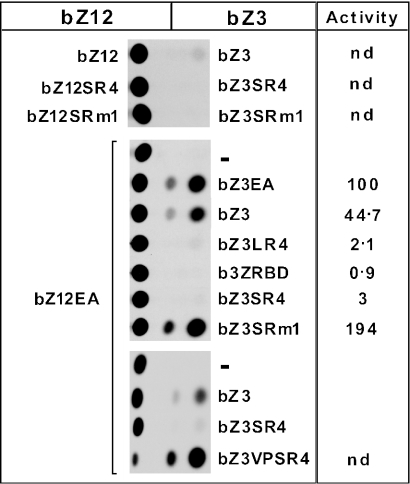
The Ewings Sarcoma Oncoprotein (EWS) interacts with several components of the mammalian transcriptional and pre-mRNA splicing machinery and is also found in the cytoplasm and even on the cell surface. The apparently diverse cellular functions of EWS are, however, not well characterized. EWS harbours a potent N-terminal transcriptional activation domain (the EAD) that is revealed in the context of oncogenic EWS-fusion proteins (EFPs) and a C-terminal RNA-binding domain (RBD) that recruits pre-mRNA splicing factors and may couple transcription and splicing. In contrast to EFPs, the presumed transcriptional role of normal EWS remains enigmatic. Here, we report that multiple RGG-boxes within the RBD are necessary and sufficient for cis-repression of the EAD and that RGG-boxes can also repress in-trans, within dimeric partners. Lys can functionally substitute for Arg, indicating that the basic nature of the Arg side chain is the critical determinant of RGG-box-mediated repression. In addition to the EAD, RGG-boxes can repress a broad range of activation domains (including those of VP16, E1a and CREB), but repression can be alleviated by the simultaneous presence of more than one activation domain. We therefore propose that a key function of RGG boxes within native EWS is to restrict promiscuous activation by the EAD while still allowing EWS to enter functional transcription complexes and participate in other transactions involving pre-mRNAs.
Figures
Similar articles
-
RGG boxes within the TET/FET family of RNA-binding proteins are functionally distinct.Transcription. 2016 Aug 7;7(4):141-51. doi: 10.1080/21541264.2016.1183071. Epub 2016 May 9. Transcription. 2016. PMID: 27159574 Free PMC article.
-
Transcriptional activation by the Ewing's sarcoma (EWS) oncogene can be cis-repressed by the EWS RNA-binding domain.J Biol Chem. 2000 Jul 28;275(30):23053-8. doi: 10.1074/jbc.M002961200. J Biol Chem. 2000. PMID: 10767297
-
Multiple aromatic side chains within a disordered structure are critical for transcription and transforming activity of EWS family oncoproteins.Proc Natl Acad Sci U S A. 2007 Jan 9;104(2):479-84. doi: 10.1073/pnas.0607007104. Epub 2007 Jan 3. Proc Natl Acad Sci U S A. 2007. PMID: 17202261 Free PMC article.
-
Molecular recognition by the EWS transcriptional activation domain.Adv Exp Med Biol. 2012;725:106-25. doi: 10.1007/978-1-4614-0659-4_7. Adv Exp Med Biol. 2012. PMID: 22399321 Review.
-
TLS, EWS and TAF15: a model for transcriptional integration of gene expression.Brief Funct Genomic Proteomic. 2006 Mar;5(1):8-14. doi: 10.1093/bfgp/ell015. Epub 2006 Feb 23. Brief Funct Genomic Proteomic. 2006. PMID: 16769671 Review.
Cited by
-
The FXR1 network acts as signaling scaffold for actomyosin remodeling.bioRxiv [Preprint]. 2024 May 25:2023.11.05.565677. doi: 10.1101/2023.11.05.565677. bioRxiv. 2024. Update in: Cell. 2024 Sep 5;187(18):5048-5063.e25. doi: 10.1016/j.cell.2024.07.015. PMID: 37961296 Free PMC article. Updated. Preprint.
-
The TET family of proteins: functions and roles in disease.J Mol Cell Biol. 2009 Dec;1(2):82-92. doi: 10.1093/jmcb/mjp025. Epub 2009 Sep 24. J Mol Cell Biol. 2009. PMID: 19783543 Free PMC article. Review.
-
Acetylation dependent translocation of EWSR1 regulates CHK2 alternative splicing in response to DNA damage.Oncogene. 2022 Jul;41(29):3694-3704. doi: 10.1038/s41388-022-02383-x. Epub 2022 Jun 22. Oncogene. 2022. PMID: 35732801
-
The FXR1 network acts as a signaling scaffold for actomyosin remodeling.Cell. 2024 Sep 5;187(18):5048-5063.e25. doi: 10.1016/j.cell.2024.07.015. Epub 2024 Aug 5. Cell. 2024. PMID: 39106863
-
EppA, a putative substrate of DdERK2, regulates cyclic AMP relay and chemotaxis in Dictyostelium discoideum.Eukaryot Cell. 2006 Jul;5(7):1136-46. doi: 10.1128/EC.00383-05. Eukaryot Cell. 2006. PMID: 16835457 Free PMC article.
References
-
- Kim J., Pelletier J. Molecular genetics of chromosome translocations involving EWS and related family members. Physiol. Genomics. 1999;1:127–138. - PubMed
-
- Kovar H., Aryee D., Zoubek A. The Ewing family of Tumors and the search for the Achilles heel. Curr. Opin. Oncol. 1999;11:275–284. - PubMed
-
- Arvand A., Denny C.T. Biology of EWS/ETS fusions in Ewing's family tumors. Oncogene. 2001;20:5747–5754. - PubMed
-
- Zucman J., Delattre O., Desmaze C., Epstein A.L., Stenman G., Speleman F., Fletchers C.D.M., Aurias A., Thomas G. EWS and ATF1 gene fusion induced by t(12:22) translocation in malignant melanoma of soft parts. Nature Genet. 1993;4:341–345. - PubMed
