

Association with class IIa histone deacetylases upregulates the sumoylation of MEF2 transcription factors
- PMID: 15743823
- PMCID: PMC1061617
- DOI: 10.1128/MCB.25.6.2273-2287.2005
Association with class IIa histone deacetylases upregulates the sumoylation of MEF2 transcription factors
Erratum in
- Mol Cell Biol. 2006 Apr;26(8):3335
Abstract
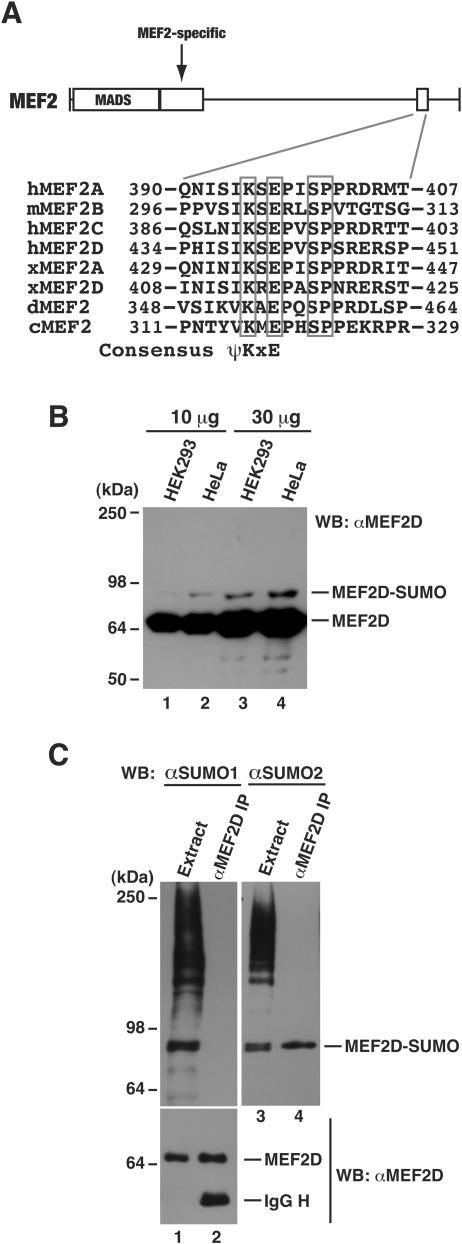
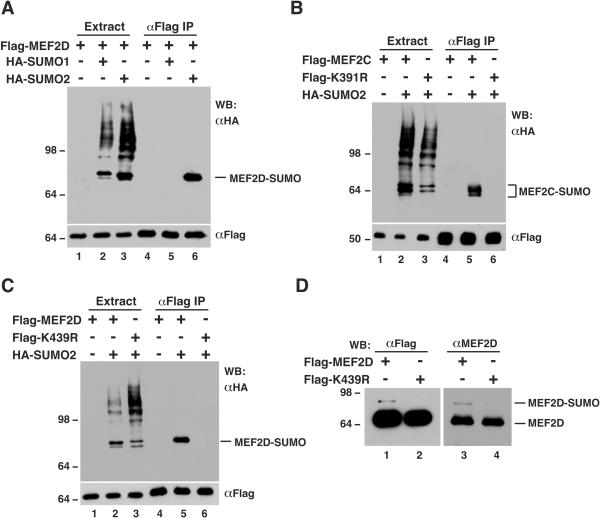
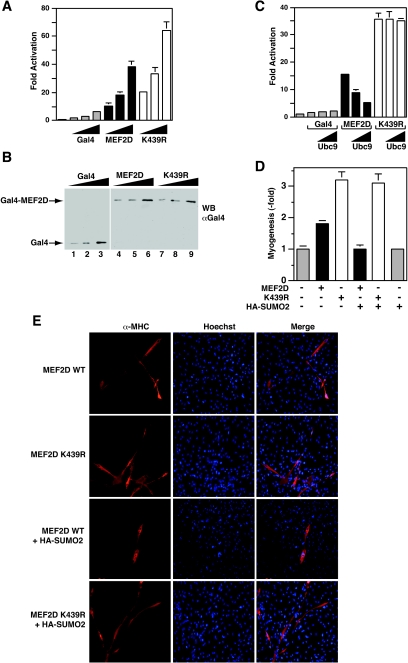
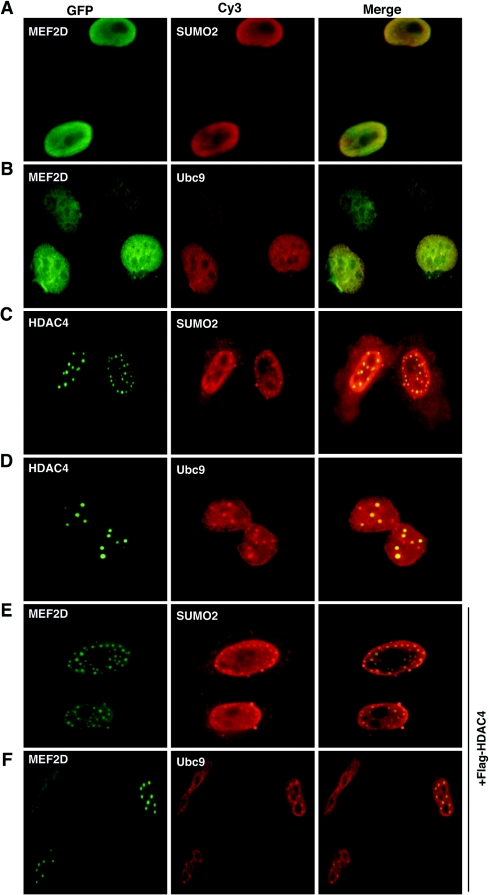
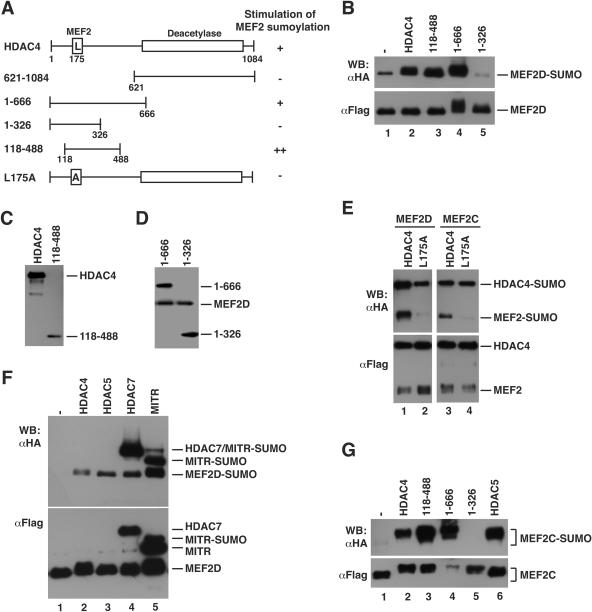
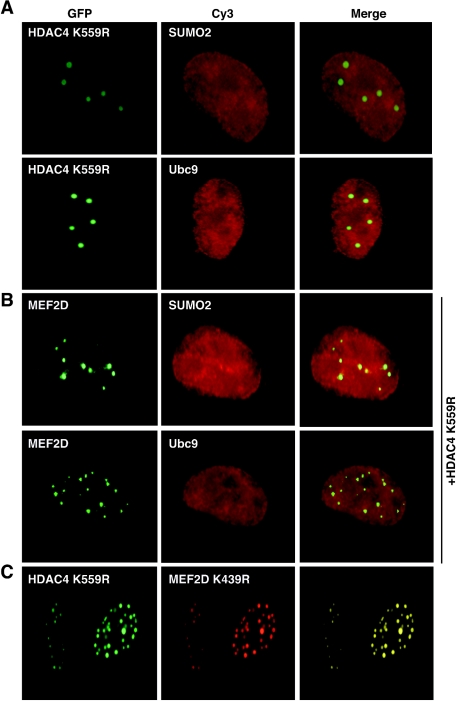
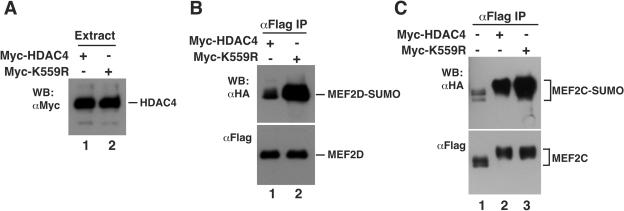
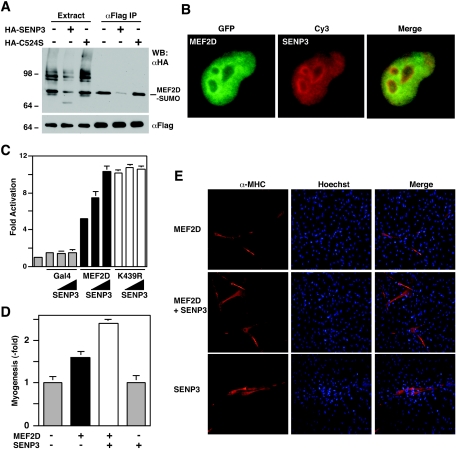
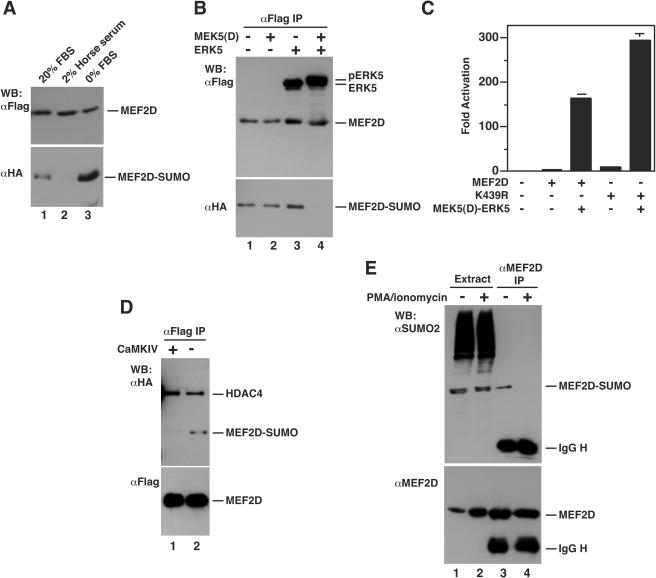
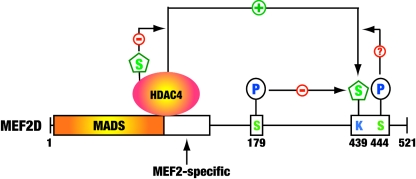
The myocyte enhancer factor-2 (MEF2) family of transcription factors plays an important role in regulating cellular programs like muscle differentiation, neuronal survival, and T-cell apoptosis. Multisite phosphorylation is known to control the transcriptional activity of MEF2 proteins, but it is unclear whether other modifications are involved. Here, we report that human MEF2D, as well as MEF2C, is modified by SUMO2 and SUMO3 at a motif highly conserved among MEF2 proteins from diverse organisms. This motif is located within the C-terminal transcriptional activation domain, and its sumoylation inhibits transcription. As a transcriptional corepressor of MEF2, histone deacetylase 4 (HDAC4) potentiates sumoylation. This potentiation is dependent on the N-terminal region but not the C-terminal deacetylase domain of HDAC4 and is inhibited by the sumoylation of HDAC4 itself. Moreover, HDAC5, HDAC7, and an HDAC9 isoform also stimulate sumoylation of MEF2. Opposing the action of class IIa deacetylases, the SUMO protease SENP3 reverses the sumoylation to augment the transcriptional and myogenic activities of MEF2. Similarly, the calcium/calmodulin-dependent kinases [corrected] and extracellular signal-regulated kinase 5 signaling pathways negatively regulate the sumoylation. These results thus identify sumoylation as a novel regulatory mechanism for MEF2 and suggest that this modification interplays with phosphorylation to promote intramolecular signaling for coordinated regulation in vivo.
Figures
Similar articles
-
Histone deacetylase 3 interacts with and deacetylates myocyte enhancer factor 2.Mol Cell Biol. 2007 Feb;27(4):1280-95. doi: 10.1128/MCB.00882-06. Epub 2006 Dec 11. Mol Cell Biol. 2007. PMID: 17158926 Free PMC article.
-
Regulation of MEF2 by histone deacetylase 4- and SIRT1 deacetylase-mediated lysine modifications.Mol Cell Biol. 2005 Oct;25(19):8456-64. doi: 10.1128/MCB.25.19.8456-8464.2005. Mol Cell Biol. 2005. PMID: 16166628 Free PMC article.
-
Control of MEF2 transcriptional activity by coordinated phosphorylation and sumoylation.J Biol Chem. 2006 Feb 17;281(7):4423-33. doi: 10.1074/jbc.M509471200. Epub 2005 Dec 15. J Biol Chem. 2006. PMID: 16356933
-
Class II histone deacetylases: structure, function, and regulation.Biochem Cell Biol. 2001;79(3):243-52. Biochem Cell Biol. 2001. PMID: 11467738 Review.
-
MEF2: a central regulator of diverse developmental programs.Development. 2007 Dec;134(23):4131-40. doi: 10.1242/dev.008367. Epub 2007 Oct 24. Development. 2007. PMID: 17959722 Review.
Cited by
-
Homeodomain-interacting protein kinase 2-dependent repression of myogenic differentiation is relieved by its caspase-mediated cleavage.Nucleic Acids Res. 2013 Jun;41(11):5731-45. doi: 10.1093/nar/gkt262. Epub 2013 Apr 24. Nucleic Acids Res. 2013. PMID: 23620283 Free PMC article.
-
An acetylation/deacetylation-SUMOylation switch through a phylogenetically conserved psiKXEP motif in the tumor suppressor HIC1 regulates transcriptional repression activity.Mol Cell Biol. 2007 Apr;27(7):2661-75. doi: 10.1128/MCB.01098-06. Epub 2007 Feb 5. Mol Cell Biol. 2007. PMID: 17283066 Free PMC article.
-
Role of SIRT1 in Modulating Acetylation of the Sarco-Endoplasmic Reticulum Ca2+-ATPase in Heart Failure.Circ Res. 2019 Apr 26;124(9):e63-e80. doi: 10.1161/CIRCRESAHA.118.313865. Circ Res. 2019. PMID: 30786847 Free PMC article.
-
Roles and post-translational regulation of cardiac class IIa histone deacetylase isoforms.J Physiol. 2015 Apr 15;593(8):1785-97. doi: 10.1113/jphysiol.2014.282442. Epub 2014 Nov 25. J Physiol. 2015. PMID: 25362149 Free PMC article. Review.
-
Evidence for a non-canonical role of HDAC5 in regulation of the cardiac Ncx1 and Bnp genes.Nucleic Acids Res. 2016 May 5;44(8):3610-7. doi: 10.1093/nar/gkv1496. Epub 2015 Dec 23. Nucleic Acids Res. 2016. PMID: 26704971 Free PMC article.
References
-
- Appella, E., and C. W. Anderson. 2001. Post-translational modifications and activation of p53 by genotoxic stresses. Eur. J. Biochem. 268:2764-2772. - PubMed
-
- Bakin, R. E., and M. O. Jung. 2004. Cytoplasmic sequestration of HDAC7 from mitochondrial and nuclear compartments upon initiation of apoptosis. J. Biol. Chem. 279:51218-51225. - PubMed
-
- Bernier-Villamor, V., D. A. Sampson, M. J. Matunis, and C. D. Lima. 2002. Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1. Cell 108:345-356. - PubMed
-
- Best, J. L., S. Ganiatsas, S. Agarwal, A. Changou, P. Salomoni, O. Shirihai, P. B. Meluh, P. P. Pandolfi, and L. I. Zon. 2002. SUMO-1 protease-1 regulates gene transcription through PML. Mol. Cell 10:843-855. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials