

The mechanism underlying the contractile effect of a chemotactic peptide, formyl-Met-Leu-Phe on the guinea-pig Taenia coli
- PMID: 15735654
- PMCID: PMC1576144
- DOI: 10.1038/sj.bjp.0706181
The mechanism underlying the contractile effect of a chemotactic peptide, formyl-Met-Leu-Phe on the guinea-pig Taenia coli
Abstract
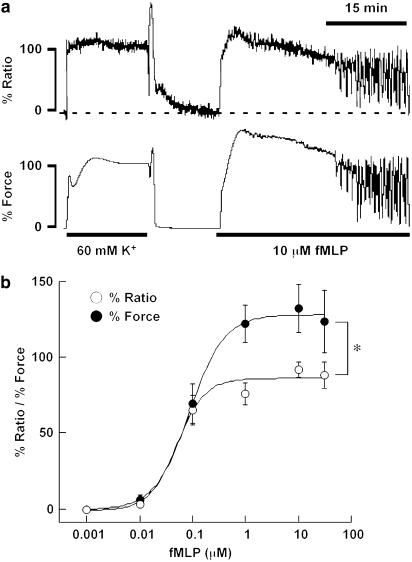
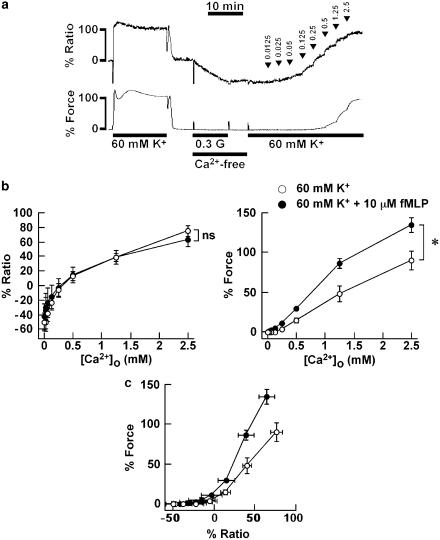
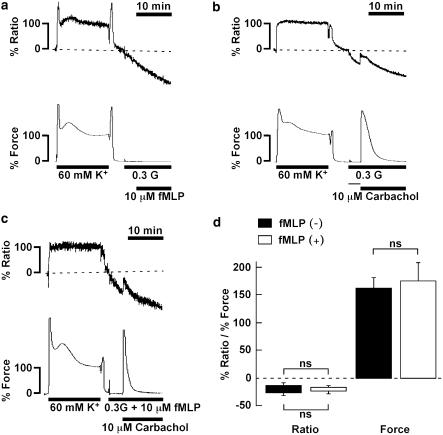
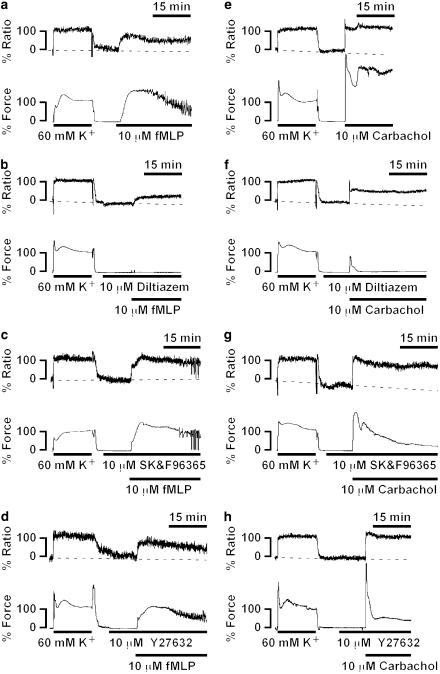
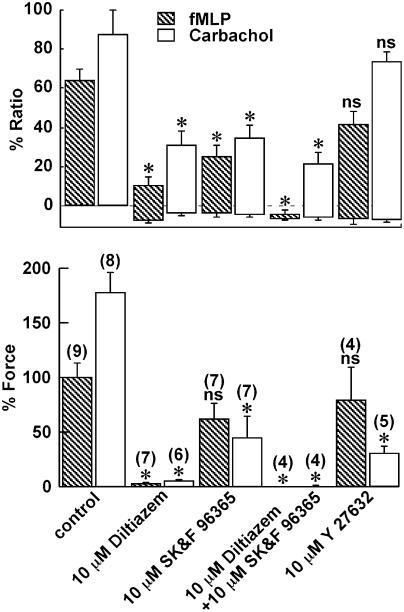
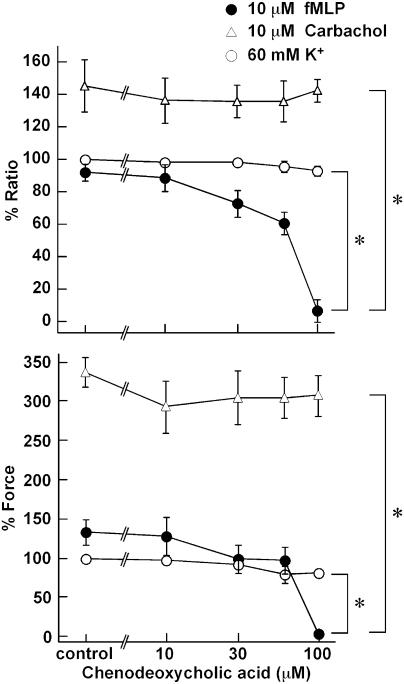
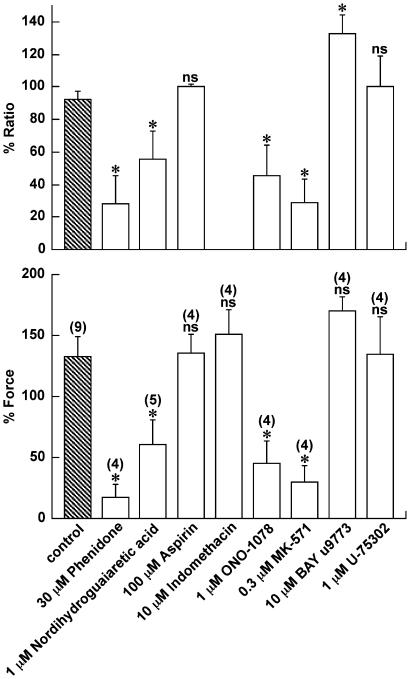
1 The contractile mechanism of N-formyl-L-methionyl-L-leucyl-L-phenylalanine (fMLP) was investigated in the guinea-pig Taenia coli, by simultaneously monitoring the changes in the cytosolic Ca(2+) concentration ([Ca(2+)](i)) and force. 2 fMLP induced a significant elevation of [Ca(2+)](i) and force at concentrations higher than 10 nM. The maximal response was obtained at a concentration of higher than 1 microM. 3 fMLP (10 microM) augmented the force development induced by a stepwise increment of the extracellular Ca(2+) concentration during 60 mM K(+) depolarization, while it had no effect on the [Ca(2+)](i) elevation, and thus produced a greater force for a given elevation of [Ca(2+)](i) than 60 mM K(+) depolarization. 4 The removal of extracellular Ca(2+) completely abolished the fMLP-induced contraction. The fMLP-induced [Ca(2+)](i) elevation was inhibited substantially but not completely by 10 microM diltiazem, partly by 10 microM SK&F 96365, and completely by their combination. 5 Y27632, a specific inhibitor of rho-kinase, had no significant effect on the fMLP-induced [Ca(2+)](i) elevation and force development. 6 Chenodeoxycholic acid, a formyl peptide receptor antagonist, specifically abolished the fMLP-induced contraction but not high K(+)- or carbachol-induced contractions. 7 A dual lipoxygenase/cyclooxygenase inhibitor, a 5-lipoxygenase inhibitor, a nonselective leukotriene receptor antagonist, and a selective type 1 cysteinyl-containing leukotriene receptor antagonist specifically reduced the fMLP-induced contraction. 8 We suggest that the low-affinity-type fMLP receptor and lipoxygenase metabolites of arachidonic acid are involved in the fMLP-induced contraction in the guinea-pig T. coli. This contraction mainly depends on the [Ca(2+)](i) elevation due to Ca(2+) influx and the enhancement of Ca(2+) sensitivity in the contractile apparatus.
Figures
Similar articles
-
Deactivation of guinea pig pulmonary alveolar macrophage responses to N-formyl-methionyl-leucyl-phenylalanine: chemotaxis, superoxide generation, and binding.J Immunol. 1985 Mar;134(3):1823-6. J Immunol. 1985. PMID: 2981922
-
Pharmacological modulation of the effects of N-formyl-L-methionyl-L-leucyl-L-phenylalanine in guinea-pigs: involvement of the arachidonic acid cascade.Br J Pharmacol. 1986 Oct;89(2):349-59. doi: 10.1111/j.1476-5381.1986.tb10267.x. Br J Pharmacol. 1986. PMID: 3096407 Free PMC article.
-
Relaxant effect of N-formyl-methionyl-leucyl-phenylalanine on rabbit vascular strips.J Pharmacol Exp Ther. 1989 Feb;248(2):774-80. J Pharmacol Exp Ther. 1989. PMID: 2537416
-
The main functions and structural modifications of tripeptide N-formyl-methionyl-leucyl-phenylalanine (fMLP) as a chemotactic factor.Pharmazie. 2008 Nov;63(11):779-83. Pharmazie. 2008. PMID: 19069235 Review.
-
Structure-activity relationship of for-L-Met L-Leu-L-Phe-OMe analogues in human neutrophils.Bioorg Chem. 2006 Oct;34(5):298-318. doi: 10.1016/j.bioorg.2006.07.001. Epub 2006 Aug 17. Bioorg Chem. 2006. PMID: 16919307 Review.
Cited by
-
Inhibition of 15-lipoxygenase (15-LOX) reverses hypoxia-induced down-regulation of potassium channels Kv1.5 and Kv2.1Inhibition of 15-lipoxygenase (15-LOX) reverses hypoxia-induced down-regulation of potassium channels Kv1.5 and Kv2.1.Int J Clin Exp Med. 2014 Nov 15;7(11):4147-53. eCollection 2014. Int J Clin Exp Med. 2014. PMID: 25550925 Free PMC article.
References
-
- AMANO M., ITO M., KIMURA K., FUKATA Y., CHIHARA K., NAKANO T., MATSUURA Y., KAIBUCHI K. Phosphorylation and activation of myosin by Rho-associated kinase (Rho-kinase) J. Biol. Chem. 1996;271:20246–20249. - PubMed
-
- ARMOUR C.L., BLACK J.L., JOHNSON P.R., VINCENC K.S., BEREND N. Formyl peptide-induced contraction of human airways in vitro. J. Appl. Physiol. 1986;60:141–146. - PubMed
-
- BAO L., GERARD N.P., EDDY R.L., JR, SHOWS T.B., GERARD C. Mapping of genes for the human C5a receptor (C5AR), human FMLP receptor (FPR), and two FMLP receptor homologue orphan receptors (FPRH1, FPRH2) to chromosome 19. Genomics. 1992;13:437–440. - PubMed
-
- BECKER E.L., FOROUHAR F.A., GRUNNET M.L., BOULAY F., TARDIF M., BORMANN B.J., SODJA D., YE R.D., WOSKA J.R., MURPHY P.M. Broad immunocytochemical localization of the formyl peptide receptor in human organs, tissues, and cells. Cell Tissue Res. 1998;292:129–135. - PubMed
-
- BOUKILI M.A., BUREAU M., LAGENTE V., LEFORT J., LELLOUCH-TUBIANA A., MALANCHERE E., VARGAFTIG B.B. Pharmacological modulation of the effects of N-formyl-L-methionyl-L-leucyl-L-phenylalanine in guinea-pigs: involvement of the arachidonic acid cascade. Br. J. Pharmacol. 1986;89:349–359. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
