

Characterization of purified intraembryonic hematopoietic stem cells as a tool to define their site of origin
- PMID: 15623562
- PMCID: PMC544043
- DOI: 10.1073/pnas.0402270102
Characterization of purified intraembryonic hematopoietic stem cells as a tool to define their site of origin
Abstract
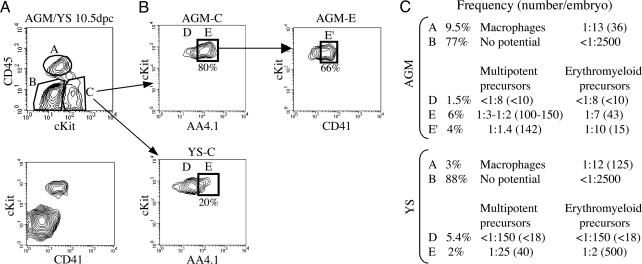
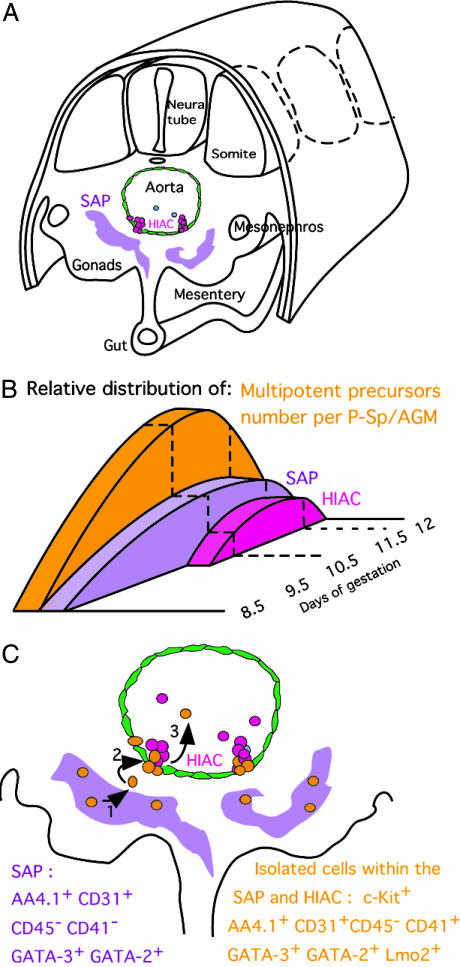
Little is known about hematopoietic stem cell (HSC) development from mesoderm. To gain more information on the intraembryonic HSC site of origin, we purified multipotent hematopoietic progenitors from the aorta-gonads-mesonephros (AGM) of mice. This population, expressing c-Kit, AA4.1, CD31, and CD41, but not Flk1, and mainly negative for CD45, proved capable of long-term reconstitution in sublethally irradiated Rag2gammac(-/-) recipients. We assigned the expression of GATA-2, GATA-3, and lmo2 to AGM-HSC, whereas erythromyeloid progenitors express only GATA-2. This unique combination of surface markers and transcription factors could be allocated in the AGM to the intraaortic clusters and the subaortic patches underlying aortic endothelial cells. Taken together, those data indicate that embryonic HSCs (i) differ from their fetal liver and adult counterpart by the low expression of CD45, (ii) do not colocalize with aortic endothelial cells as previously thought, and (iii) are localized, at 10.5 days postcoitum, in the splanchnic mesoderm underlying aortic endothelial cells, within GATA-3(+)CD31(+) cell clusters.
Figures



Similar articles
-
Progressive divergence of definitive haematopoietic stem cells from the endothelial compartment does not depend on contact with the foetal liver.Development. 2005 Sep;132(18):4179-91. doi: 10.1242/dev.01974. Epub 2005 Aug 17. Development. 2005. PMID: 16107475
-
Expansive effects of aorta-gonad-mesonephros-derived stromal cells on hematopoietic stem cells from embryonic stem cells.Chin Med J (Engl). 2005 Dec 5;118(23):1979-86. Chin Med J (Engl). 2005. PMID: 16336834
-
CD45(low)c-Kit(high) cells have hematopoietic properties in the mouse aorta-gonad-mesonephros region.Exp Cell Res. 2012 Apr 1;318(6):705-15. doi: 10.1016/j.yexcr.2012.01.017. Epub 2012 Jan 25. Exp Cell Res. 2012. PMID: 22289156
-
The embryonic aorta-gonad-mesonephros region as a generator of haematopoietic stem cells.APMIS. 2005 Nov-Dec;113(11-12):804-12. doi: 10.1111/j.1600-0463.2005.apm_368.x. APMIS. 2005. PMID: 16480451 Review.
-
Genetic and functional characterization of isolated stromal cell lines from the aorta-gonado-mesonephros region.Ann N Y Acad Sci. 2005 Jun;1044:51-9. doi: 10.1196/annals.1349.007. Ann N Y Acad Sci. 2005. PMID: 15958697 Review.
Cited by
-
Megakaryocyte production is sustained by direct differentiation from erythromyeloid progenitors in the yolk sac until midgestation.Immunity. 2021 Jul 13;54(7):1433-1446.e5. doi: 10.1016/j.immuni.2021.04.026. Epub 2021 May 31. Immunity. 2021. PMID: 34062116 Free PMC article.
-
Specification of tissue-resident macrophages during organogenesis.Science. 2016 Sep 9;353(6304):aaf4238. doi: 10.1126/science.aaf4238. Epub 2016 Aug 4. Science. 2016. PMID: 27492475 Free PMC article.
-
Definitive hematopoiesis initiates through a committed erythromyeloid progenitor in the zebrafish embryo.Development. 2007 Dec;134(23):4147-56. doi: 10.1242/dev.012385. Epub 2007 Oct 24. Development. 2007. PMID: 17959717 Free PMC article.
-
Rac1 is essential for intraembryonic hematopoiesis and for the initial seeding of fetal liver with definitive hematopoietic progenitor cells.Blood. 2008 Apr 1;111(7):3313-21. doi: 10.1182/blood-2007-08-110114. Epub 2007 Dec 14. Blood. 2008. PMID: 18083846 Free PMC article.
-
Gastruloids as in vitro models of embryonic blood development with spatial and temporal resolution.Sci Rep. 2022 Aug 4;12(1):13380. doi: 10.1038/s41598-022-17265-1. Sci Rep. 2022. PMID: 35927563 Free PMC article.
References
-
- Cumano, A., Dieterlen-Lievre, F. & Godin, I. (1996) Cell 86, 907-916. - PubMed
-
- Cumano, A., Ferraz, J. C., Klaine, M., Di Santo, J. P. & Godin, I. (2001) Immunity 15, 477-485. - PubMed
-
- Muller, A. M., Medvinsky, A., Strouboulis, J., Grosveld, F. & Dzierzak, E. (1994) Immunity 1, 291-301. - PubMed
-
- Yoder, M. C., Hiatt, K., Dutt, P., Mukherjee, P., Bodine, D. M. & Orlic, D. (1997) Immunity 7, 335-344. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
