

MEKK1 transduces activin signals in keratinocytes to induce actin stress fiber formation and migration
- PMID: 15601830
- PMCID: PMC538759
- DOI: 10.1128/MCB.25.1.60-65.2005
MEKK1 transduces activin signals in keratinocytes to induce actin stress fiber formation and migration
Abstract
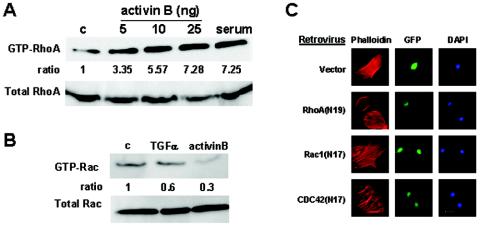
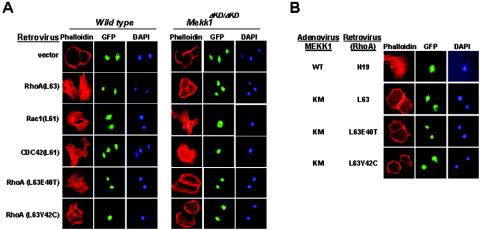
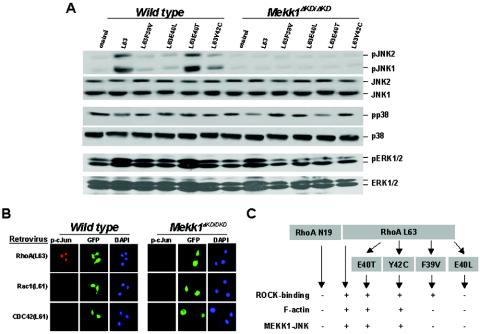
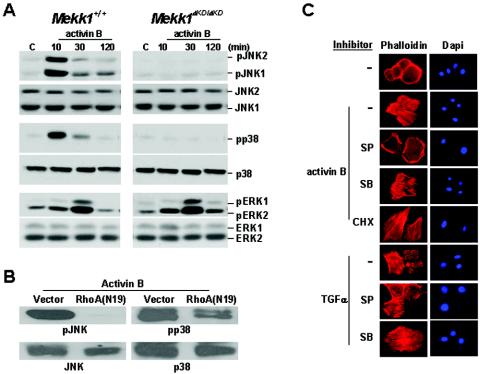
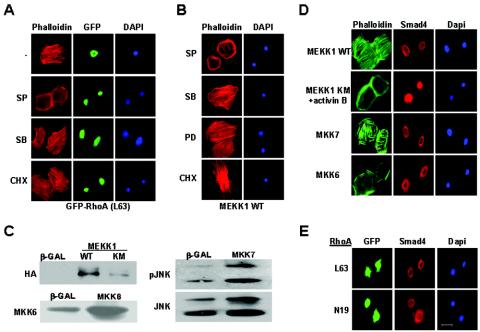
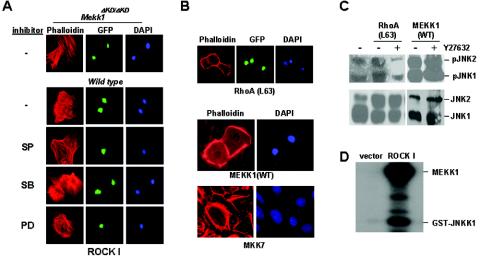
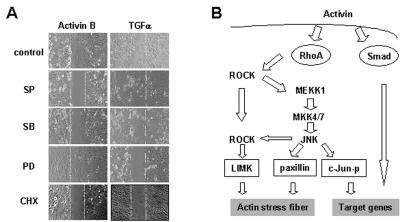
Activins and other members of the transforming growth factor beta family play a critical role in morphological changes of the epidermis that require epithelial cell movement. We investigated the molecular pathways in the transmission of activin signals that lead to actin reorganization and epithelial cell migration. We found that activins cause the activation of RhoA but not of Rac and CDC42, leading to MEKK1-dependent phosphorylation of JNK and transcription factor c-Jun. Through a RhoA-independent mechanism, the activins also induce p38 activity in keratinocytes from wild-type but not from MEKK1-deficient mice. Although neither pathway is dependent on Smad activation, the MEKK1-mediated JNK and p38 activities are both essential for activin-stimulated and transcription-dependent keratinocyte migration. Only JNK is involved in transcription-independent actin stress fiber formation, which needs also the activity of ROCK. Because ROCK is required for JNK activation by RhoA and its overexpression leads to MEKK1 activation, we propose a RhoA-ROCK-MEKK1-JNK pathway and a MEKK1-p38 pathway as Smad-independent mechanisms in the transmission of activin signals. Together, these pathways lead to the control of actin cytoskeleton reorganization and epithelial cell migration, contributing to the physiologic and pathological effects of activins on epithelial morphogenesis.
Figures
Similar articles
-
Activin B promotes epithelial wound healing in vivo through RhoA-JNK signaling pathway.PLoS One. 2011;6(9):e25143. doi: 10.1371/journal.pone.0025143. Epub 2011 Sep 19. PLoS One. 2011. PMID: 21949871 Free PMC article.
-
The signaling pathways in tissue morphogenesis: a lesson from mice with eye-open at birth phenotype.Biochem Pharmacol. 2004 Sep 15;68(6):997-1001. doi: 10.1016/j.bcp.2004.05.028. Biochem Pharmacol. 2004. PMID: 15313393
-
A role for MEK kinase 1 in TGF-beta/activin-induced epithelium movement and embryonic eyelid closure.EMBO J. 2003 Sep 1;22(17):4443-54. doi: 10.1093/emboj/cdg440. EMBO J. 2003. PMID: 12941696 Free PMC article.
-
[The Rho protein family and its role in the cellular cytoskeleton].Postepy Hig Med Dosw (Online). 2008 Mar 10;62:110-7. Postepy Hig Med Dosw (Online). 2008. PMID: 18334926 Review. Polish.
-
Sticky business: orchestrating cellular signals at adherens junctions.Cell. 2003 Feb 21;112(4):535-48. doi: 10.1016/s0092-8674(03)00108-9. Cell. 2003. PMID: 12600316 Review.
Cited by
-
Genetic Control of MAP3K1 in Eye Development and Sex Differentiation.Cells. 2021 Dec 23;11(1):34. doi: 10.3390/cells11010034. Cells. 2021. PMID: 35011600 Free PMC article. Review.
-
The Epstein-Barr virus encoded LMP1 oncoprotein modulates cell adhesion via regulation of activin A/TGFβ and β1 integrin signalling.Sci Rep. 2016 Jan 19;6:19533. doi: 10.1038/srep19533. Sci Rep. 2016. PMID: 26782058 Free PMC article.
-
Calponin-3 deficiency augments contractile activity, plasticity, fibrogenic response and Yap/Taz transcriptional activation in lens epithelial cells and explants.Sci Rep. 2020 Jan 28;10(1):1295. doi: 10.1038/s41598-020-58189-y. Sci Rep. 2020. PMID: 31992794 Free PMC article.
-
Activins and Inhibins: Roles in Development, Physiology, and Disease.Cold Spring Harb Perspect Biol. 2016 Jul 1;8(7):a021881. doi: 10.1101/cshperspect.a021881. Cold Spring Harb Perspect Biol. 2016. PMID: 27328872 Free PMC article. Review.
-
BMP2 and BMP7 play antagonistic roles in feather induction.Development. 2008 Aug;135(16):2797-805. doi: 10.1242/dev.018341. Epub 2008 Jul 17. Development. 2008. PMID: 18635609 Free PMC article.
References
-
- Ashcroft, G. S., X. Yang, A. B. Glick, M. Weinstein, J. L. Letterio, D. E. Mizel, M. Anzano, T. Greenwell-Wild, S. M. Wahl, C. Deng, and A. B. Roberts. 1999. Mice lacking Smad3 show accelerated wound healing and an impaired local inflammatory response. Nat. Cell Biol. 1:260-266. - PubMed
-
- Gallagher, E. D., S. Gutowski, P. C. Sternweis, and M. H. Cobb. 2003. RhoA binds to the amino terminus of MEKK1 and regulates its kinase activity. J. Biol. Chem. - PubMed
-
- Hall, A. 1998. Rho GTPases and the actin cytoskeleton. Science 279:509-514. - PubMed
-
- Huang, C., Z. Rajfur, C. Borchers, M. D. Schaller, and K. Jacobson. 2003. JNK phosphorylates paxillin and regulates cell migration. Nature 424:219-223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous