

Comparative Study
doi: 10.1101/gad.1255404.
Epub 2004 Dec 1.
The AMP-activated protein kinase AAK-2 links energy levels and insulin-like signals to lifespan in C. elegans
Affiliations
- PMID: 15574588
- PMCID: PMC535911
- DOI: 10.1101/gad.1255404
Item in Clipboard
Comparative Study
The AMP-activated protein kinase AAK-2 links energy levels and insulin-like signals to lifespan in C. elegans
Genes Dev.
.
Erratum in
- Genes Dev. 2005 Feb 1;19(3):411
- Genes Dev. 2005 Jan 1;19(1):188
Abstract
Although limiting energy availability extends lifespan in many organisms, it is not understood how lifespan is coupled to energy levels. We find that the AMP:ATP ratio, a measure of energy levels, increases with age in Caenorhabditis elegans and can be used to predict life expectancy. The C. elegans AMP-activated protein kinase alpha subunit AAK-2 is activated by AMP and functions to extend lifespan. In addition, either an environmental stressor that increases the AMP:ATP ratio or mutations that lower insulin-like signaling extend lifespan in an aak-2-dependent manner. Thus, AAK-2 is a sensor that couples lifespan to information about energy levels and insulin-like signals.
Figures
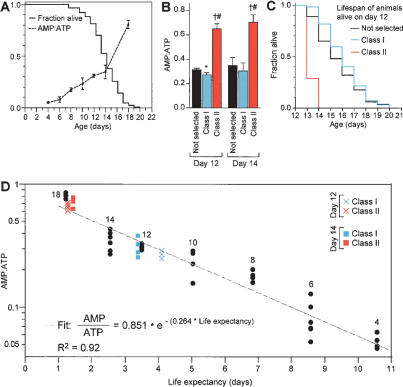
The AMP:ATP ratio predicts life expectancy. (A) Survival curve and increasing AMP:ATP ratio with age. fem-1(hc17); fer-15(b26) animals were grown at 25°C; their lack of sperm at this temperature facilitates growing a large number of animals. In parallel cultures, either the number of surviving animals or adenine nucleotide measurements were recorded at the times indicated. (B) AMP:ATP ratios of 12- and 14-d animals divided based on their motility. “Not selected” represents a group of animals that was not separated based on motility. ★P < 0.01 and †P < 0.0001, compared with “not selected”; #P < 0.0001 compared with Class I (unpaired t-test). (C) Survival curves of 12-d Class I or Class II or “not selected” animals based on motility. Similar data were obtained for 14-d animals. For statistical data on lifespans see Supplementary Table 1. (D) Least squares regression of the AMP:ATP ratio as a function of life expectancy of animals described in A. The age of “not selected” animals is noted next to the data points. The fit and R2 values were determined from “not selected” animals only. Twelve- and fourteen-day Class I and Class II animals were plotted on this graph according to AMP:ATP ratio and life expectancy (measured in B,C); actual life expectancy closely correlated with values predicted based on AMP:ATP ratio. The increase in the AMP:ATP ratio with age is not due solely to an increase in the fraction of Class II relative to Class I animals, because the AMP:ATP ratio of day-12 Class I animals is higher than that of day-4 animals, which are all Class I (P < 0.0001, unpaired t-test).
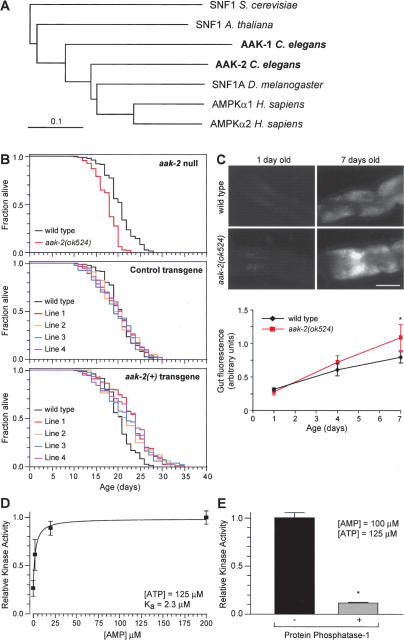
AAK-2 has kinase activity and functions to extend lifespan. (A) Phylogenetic analysis of the AAK-1 and AAK-2 sequences. Additional sequences used were hAMPKα1 (GI:20178277), hAMPKα2 (GI:20178276), DmSNF1A (GI:17137472), AtSNF1 (GI:30678280), and ScSNF1 (GI:6320685). Bar is 0.1 substitutions per site. (B) Survival curves of animals with decreased, normal, or increased aak-2 gene dosage. For statistical data see Supplementary Table 1. (C) Representative pictures and quantification of fluorescence in the anterior intestine as a function of adult age and genotype (400-msec exposures; bar is 50 μm; anterior is left, dorsal is up). ★P < 0.0001; otherwise P > 0.05 (unpaired t-test). (D,E) Measurements of kinase activity in AAK-2-GFP immunoprecipitates using the SAMS peptide assay. (D) Increasing the AMP concentration leads to an increase in kinase activity. (E) Treatment with protein phosphatase-1 greatly reduces kinase activity. ★P < 0.0001 compared with no treatment (unpaired t-test).
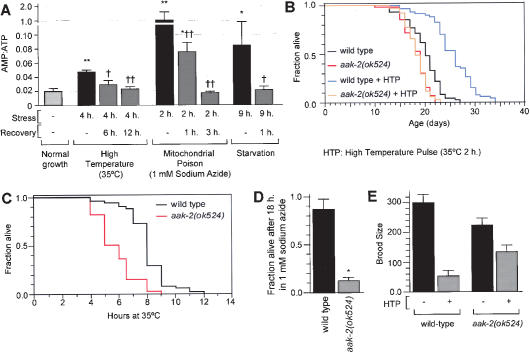
Effect of stress on wild-type and aak-2 mutants. (A) AMP:ATP ratio in wild-type animals under normal and stressful conditions. Comparison of stress condition with normal growth (★), and comparison of stress condition with reversal of stress (†). ★ and †, P < 0.01; ★★ and ††, P < 0.0001 (ANOVA). (B) Effect of a high-temperature pulse on the lifespans of wild-type animals and aak-2(ok524) mutants. (HTP) Pre-fertile 1-h-old adults exposed for 2 h to 35°C. For statistical data see Supplementary Table 1. (C) Survival of animals cultured at 35°C. aak-2(ok524) animals have 28% shorter lifespans than wild type (P < 0.0001, logrank test). aak-2(RNAi) animals also have short lifespans at 35°C compared with control animals (data not shown). (D) Sensitivity to killing by 1 mM sodium azide for 18 h. ★P < 0.0001 compared with wild type (unpaired t-test). No untreated control animals of either genotype died after 18 h (data not shown). (E) Effect of a high-temperature pulse (2 h for 35°C) on the brood sizes of wild-type animals and aak-2(ok524) mutants. P < 0.0001 for all pair-wise comparisons (ANOVA).
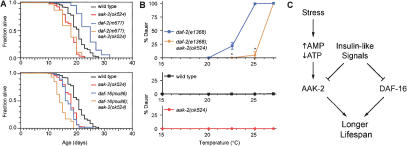
Interactions between aak-2 and insulin-like pathway mutants. (A) Effect of aak-2(ok524) on the lifespans of daf-2 and daf-16 mutants. For statistical data see Supplementary Table 1. (B) Role of aak-2 on dauer formation. ★P < 0.0001 compared with daf-2(e1368) (ANOVA). Bottom panels show that wild-type animals and aak-2(ok524) mutants do not form dauers at the temperatures tested. aak-2(RNAi) also suppresses the dauer-constitutive and lifespan extension phenotypes of daf-2(e1368) mutants (data not shown). (C) Model. AAK-2 regulates lifespan in response to changes in the AMP:ATP ratio and insulin-like signals.
Similar articles
-
Aging networks in Caenorhabditis elegans: AMP-activated protein kinase (aak-2) links multiple aging and metabolism pathways.Aging Cell. 2006 Apr;5(2):119-26. doi: 10.1111/j.1474-9726.2006.00205.x. Aging Cell. 2006. PMID: 16626391
-
5'-AMP-activated protein kinase signaling in Caenorhabditis elegans.Exp Biol Med (Maywood). 2008 Jan;233(1):12-20. doi: 10.3181/0705-MR-117. Exp Biol Med (Maywood). 2008. PMID: 18156301 Review.
-
Chronic corticosterone injections induce a decrease of ATP levels and sustained activation of AMP-activated protein kinase in hippocampal tissues of male mice.Brain Res. 2008 Jan 29;1191:148-56. doi: 10.1016/j.brainres.2007.11.027. Epub 2007 Nov 26. Brain Res. 2008. PMID: 18164281
-
Modulation of the activities of AMP-activated protein kinase, protein kinase B, and mammalian target of rapamycin by limiting energy availability with 2-deoxyglucose.Mol Carcinog. 2008 Aug;47(8):616-28. doi: 10.1002/mc.20425. Mol Carcinog. 2008. PMID: 18247380
-
The AMP-activated protein kinase: more than an energy sensor.Essays Biochem. 2007;43:121-37. doi: 10.1042/BSE0430121. Essays Biochem. 2007. PMID: 17705797 Review.
Cited by
-
JNK3 perpetuates metabolic stress induced by Aβ peptides.Neuron. 2012 Sep 6;75(5):824-37. doi: 10.1016/j.neuron.2012.06.024. Neuron. 2012. PMID: 22958823 Free PMC article.
-
Mitochondrial deficiency: a double-edged sword for aging and neurodegeneration.Front Genet. 2012 Nov 26;3:244. doi: 10.3389/fgene.2012.00244. eCollection 2012. Front Genet. 2012. PMID: 23248639 Free PMC article.
-
Zinc Levels Modulate Lifespan through Multiple Longevity Pathways in Caenorhabditis elegans.PLoS One. 2016 Apr 14;11(4):e0153513. doi: 10.1371/journal.pone.0153513. eCollection 2016. PLoS One. 2016. PMID: 27078872 Free PMC article.
-
Role of AMP-activated protein kinase in metabolic depression in animals.J Comp Physiol B. 2016 Jan;186(1):1-16. doi: 10.1007/s00360-015-0920-x. Epub 2015 Jul 15. J Comp Physiol B. 2016. PMID: 26174210 Review.
-
Enhanced energy metabolism contributes to the extended life span of calorie-restricted Caenorhabditis elegans.J Biol Chem. 2012 Sep 7;287(37):31414-26. doi: 10.1074/jbc.M112.377275. Epub 2012 Jul 18. J Biol Chem. 2012. PMID: 22810224 Free PMC article.
References
-
- Andersson U., Filipsson, K., Abbott, C.R., Woods, A., Smith, K., Bloom, S.R., Carling, D., and Small, C.J. 2004. AMP-activated protein kinase plays a role in the control of food intake. J. Biol. Chem. 279: 12005-12008. - PubMed
-
- Apfeld J. and Kenyon, C. 1999. Regulation of lifespan by sensory perception in Caenorhabditis elegans. Nature 402: 804-809. - PubMed
-
- Beauloye C., Marsin, A.S., Bertrand, L., Krause, U., Hardie, D.G., Vanoverschelde, J.L., and Hue, L. 2001. Insulin antagonizes AMP-activated protein kinase activation by ischemia or anoxia in rat hearts, without affecting total adenine nucleotides. FEBS Lett. 505: 348-352. - PubMed
-
- Cypser J.R. and Johnson, T.E. 2002. Multiple stressors in Caenorhabditis elegans induce stress hormesis and extended longevity. J. Gerontol. A Biol. Sci. Med. Sci. 57: B109-B114. - PubMed
-
- Davies S.P., Carling, D., and Hardie, D.G. 1989. Tissue distribution of the AMP-activated protein kinase, and lack of activation by cyclic-AMP-dependent protein kinase, studied using a specific and sensitive peptide assay. Eur. J. Biochem. 186: 123-128. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases