

Insufficient production and tissue delivery of CD4+ memory T cells in rapidly progressive simian immunodeficiency virus infection
- PMID: 15545355
- PMCID: PMC2211921
- DOI: 10.1084/jem.20041049
Insufficient production and tissue delivery of CD4+ memory T cells in rapidly progressive simian immunodeficiency virus infection
Abstract
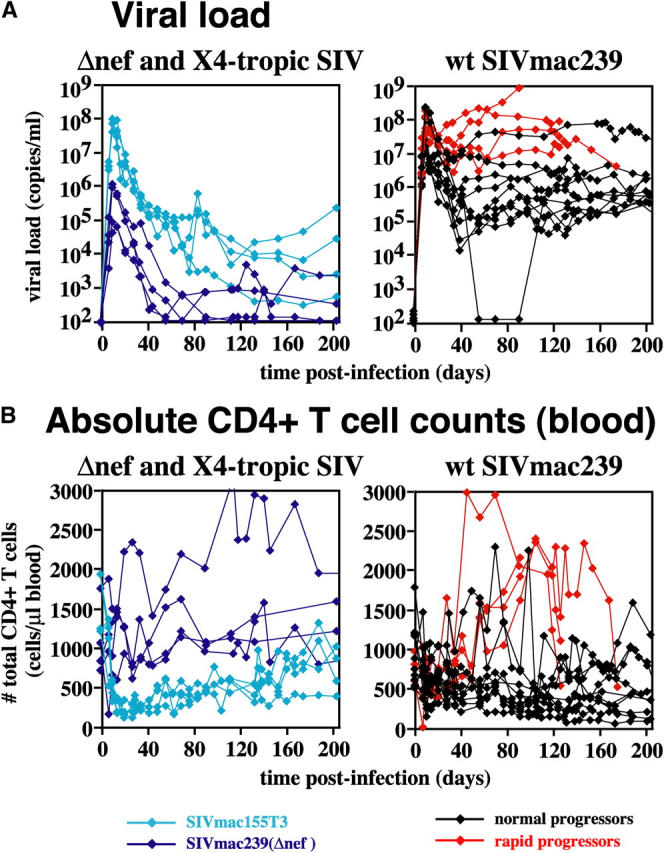
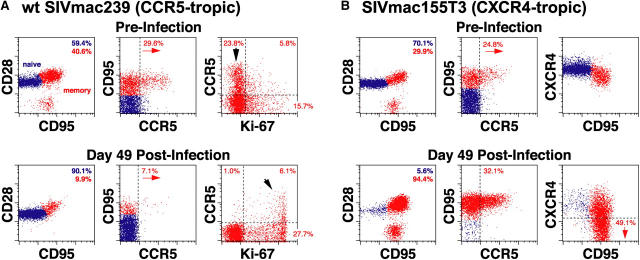
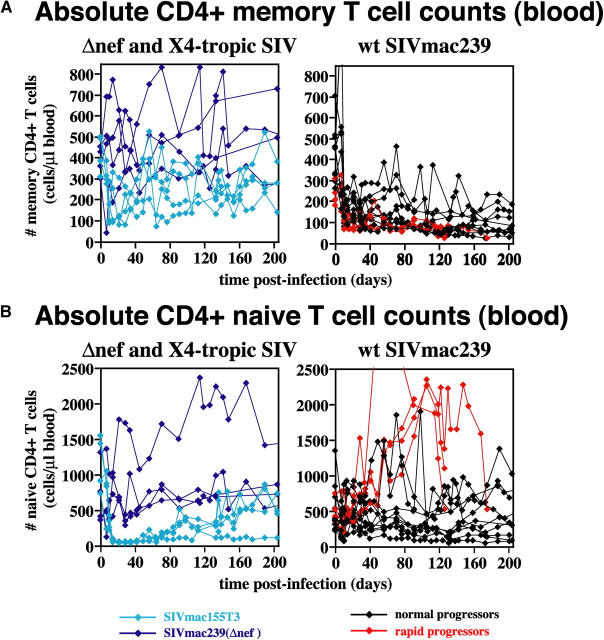
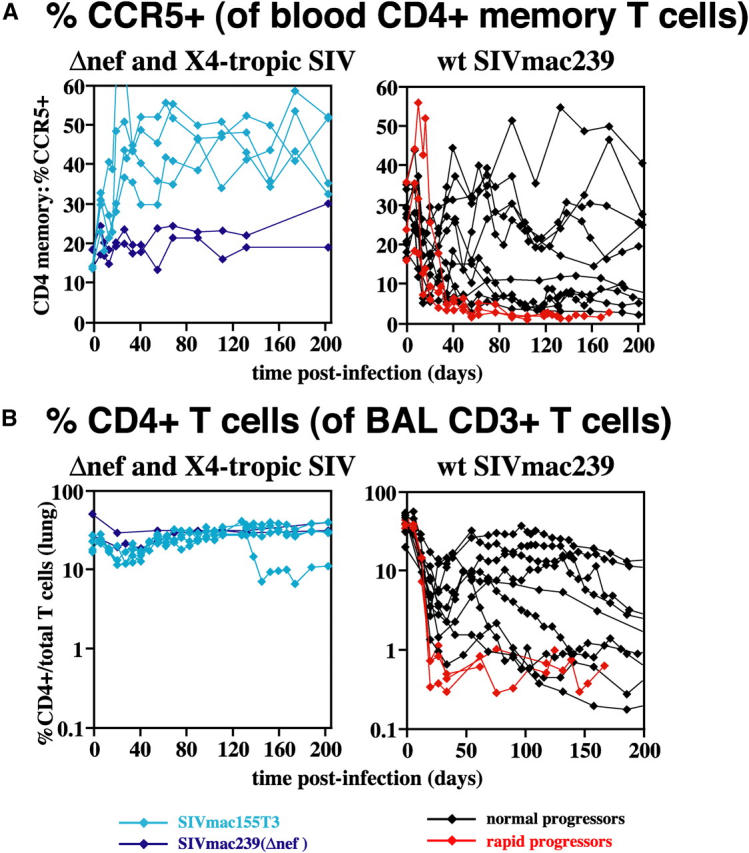
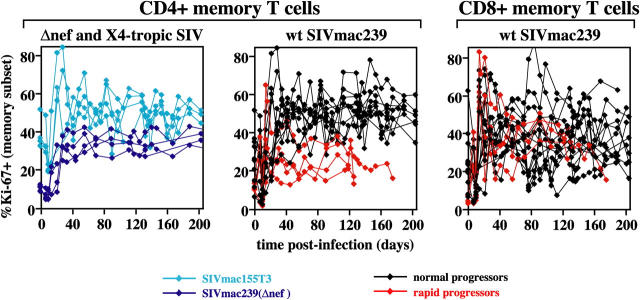
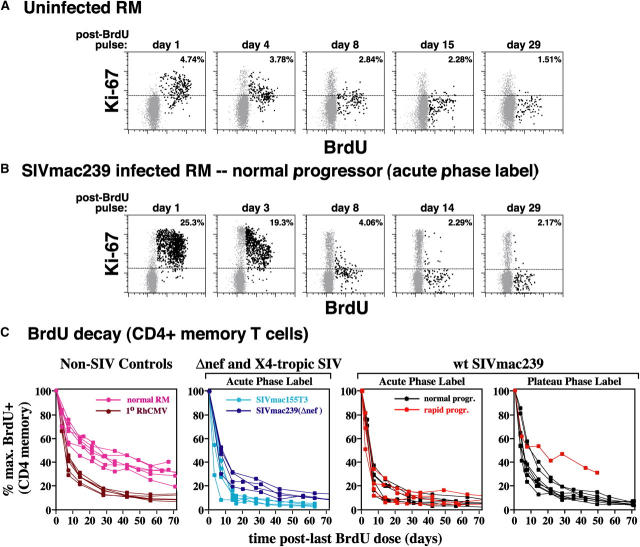
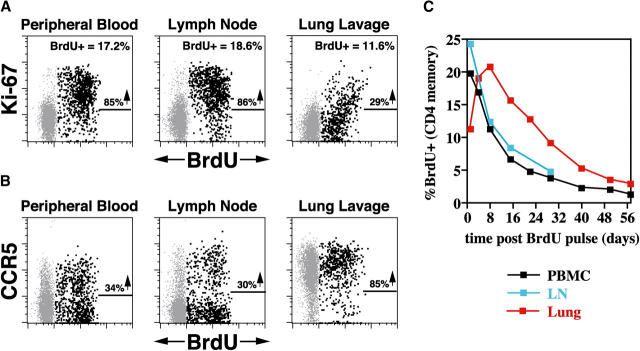
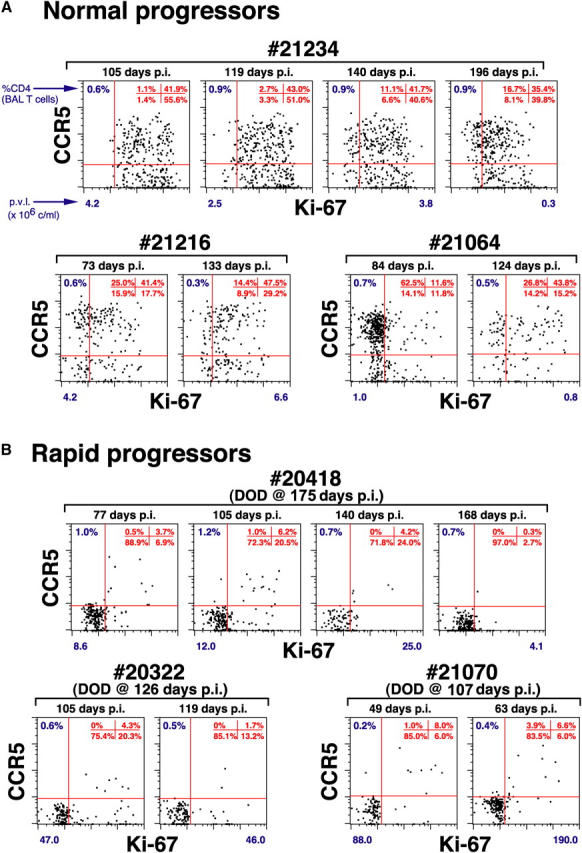
The mechanisms linking human immunodeficiency virus replication to the progressive immunodeficiency of acquired immune deficiency syndrome are controversial, particularly the relative contribution of CD4+ T cell destruction. Here, we used the simian immunodeficiency virus (SIV) model to investigate the relationship between systemic CD4+ T cell dynamics and rapid disease progression. Of 18 rhesus macaques (RMs) infected with CCR5-tropic SIVmac239 (n=14) or CXCR4-tropic SIVmac155T3 (n=4), 4 of the former group manifested end-stage SIV disease by 200 d after infection. In SIVmac155T3 infections, naive CD4+ T cells were dramatically depleted, but this population was spared by SIVmac239, even in rapid progressors. In contrast, all SIVmac239-infected RMs demonstrated substantial systemic depletion of CD4+ memory T cells by day 28 after infection. Surprisingly, the extent of CD4+ memory T cell depletion was not, by itself, a strong predictor of rapid progression. However, in all RMs destined for stable infection, this depletion was countered by a striking increase in production of short-lived CD4+ memory T cells, many of which rapidly migrated to tissue. In all rapid progressors (P <0.0001), production of these cells initiated but failed by day 42 of infection, and tissue delivery of new CD4+ memory T cells ceased. Thus, although profound depletion of tissue CD4+ memory T cells appeared to be a prerequisite for early pathogenesis, it was the inability to respond to this depletion with sustained production of tissue-homing CD4+ memory T cells that best distinguished rapid progressors, suggesting that mechanisms of the CD4+ memory T cell generation play a crucial role in maintaining immune homeostasis in stable SIV infection.
Figures
Similar articles
-
CD4+ CCR5+ T-cell dynamics during simian immunodeficiency virus infection of Chinese rhesus macaques.J Virol. 2007 Dec;81(24):13865-75. doi: 10.1128/JVI.00452-07. Epub 2007 Sep 26. J Virol. 2007. PMID: 17898067 Free PMC article.
-
Loss of naïve cells accompanies memory CD4+ T-cell depletion during long-term progression to AIDS in Simian immunodeficiency virus-infected macaques.J Virol. 2007 Jan;81(2):893-902. doi: 10.1128/JVI.01635-06. Epub 2006 Nov 8. J Virol. 2007. PMID: 17093193 Free PMC article.
-
Differential Impact of In Vivo CD8+ T Lymphocyte Depletion in Controller versus Progressor Simian Immunodeficiency Virus-Infected Macaques.J Virol. 2015 Sep;89(17):8677-86. doi: 10.1128/JVI.00869-15. Epub 2015 Jun 10. J Virol. 2015. PMID: 26063417 Free PMC article.
-
CD4+ T-cell depletion in HIV infection: killed by friendly fire?Curr HIV/AIDS Rep. 2005 Feb;2(1):16-23. doi: 10.1007/s11904-996-0004-3. Curr HIV/AIDS Rep. 2005. PMID: 16091244 Review.
-
CD4(+) T-cell depletion in HIV infection: mechanisms of immunological failure.Immunol Rev. 2013 Jul;254(1):54-64. doi: 10.1111/imr.12066. Immunol Rev. 2013. PMID: 23772614 Free PMC article. Review.
Cited by
-
Mucosal immunity in human and simian immunodeficiency lentivirus infections.Mucosal Immunol. 2013 Jul;6(4):657-65. doi: 10.1038/mi.2013.15. Epub 2013 Apr 3. Mucosal Immunol. 2013. PMID: 23549448 Free PMC article. Review.
-
Effect of Anti-IL-15 Administration on T Cell and NK Cell Homeostasis in Rhesus Macaques.J Immunol. 2016 Aug 15;197(4):1183-98. doi: 10.4049/jimmunol.1600065. Epub 2016 Jul 18. J Immunol. 2016. PMID: 27430715 Free PMC article.
-
Virus-specific T cell responses in macaques acutely infected with SHIV(sf162p3).Virology. 2007 Jun 20;363(1):36-47. doi: 10.1016/j.virol.2007.01.010. Epub 2007 Feb 16. Virology. 2007. PMID: 17307212 Free PMC article.
-
Global genomic analysis reveals rapid control of a robust innate response in SIV-infected sooty mangabeys.J Clin Invest. 2009 Dec;119(12):3556-72. doi: 10.1172/JCI40115. J Clin Invest. 2009. PMID: 19959874 Free PMC article.
-
Simian immunodeficiency virus SIVagm dynamics in African green monkeys.J Virol. 2008 Apr;82(7):3713-24. doi: 10.1128/JVI.02402-07. Epub 2008 Jan 23. J Virol. 2008. PMID: 18216122 Free PMC article.
References
-
- Cohen, O.J., and A.S. Fauci. 2001. Pathogenesis and medical aspests of HIV-1 infection. Fields Virology. D.M. Knipe and P.M. Howley, editors. Lippincott Williams & Wilkins, Philadelphia. 2043–2094.
-
- McCune, J.M. 2001. The dynamics of CD4+ T-cell depletion in HIV disease. Nature. 410:974–979. - PubMed
-
- Douek, D.C., L.J. Picker, and R.A. Koup. 2003. T cell dynamics in HIV-1 infection. Annu. Rev. Immunol. 21:265–304. - PubMed
-
- Hazenberg, M.D., D. Hamann, H. Schuitemaker, and F. Miedema. 2000. T cell depletion in HIV-1 infection: how CD4+ T cells go out of stock. Nat. Immunol. 1:285–289. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
