

A cyclooxygenase-2 homologue encoded by rhesus cytomegalovirus is a determinant for endothelial cell tropism
- PMID: 15507640
- PMCID: PMC525102
- DOI: 10.1128/JVI.78.22.12529-12536.2004
A cyclooxygenase-2 homologue encoded by rhesus cytomegalovirus is a determinant for endothelial cell tropism
Abstract
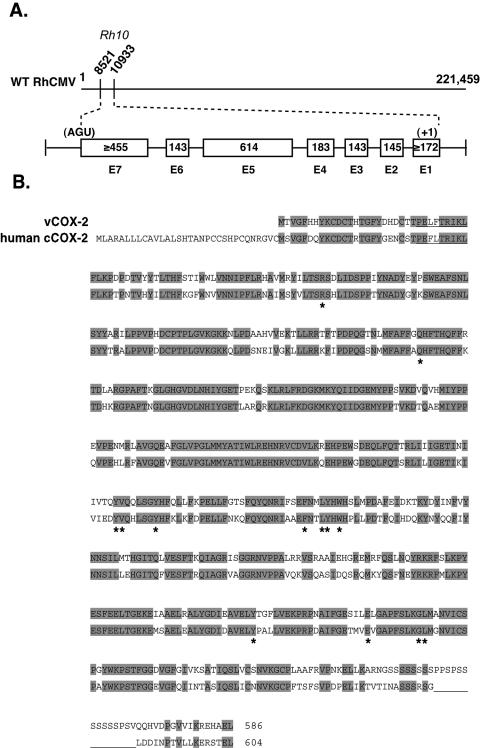
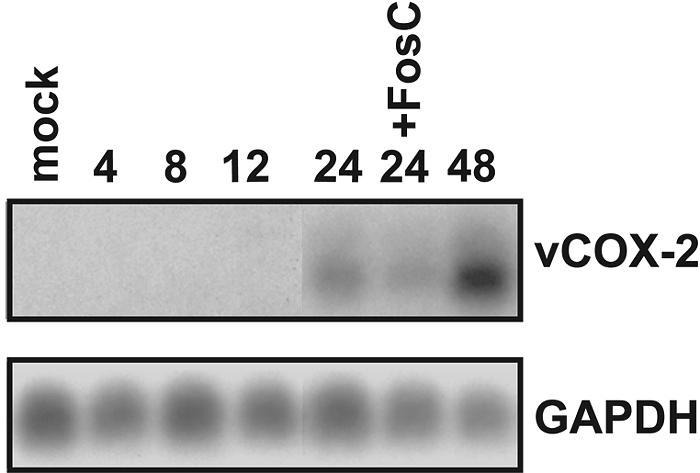
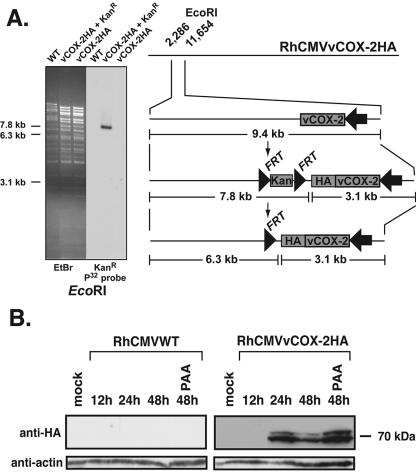
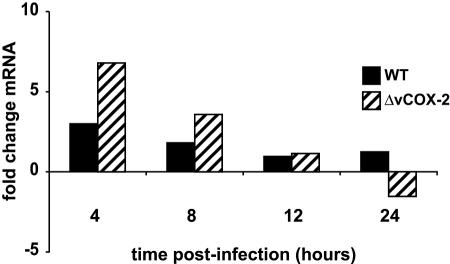
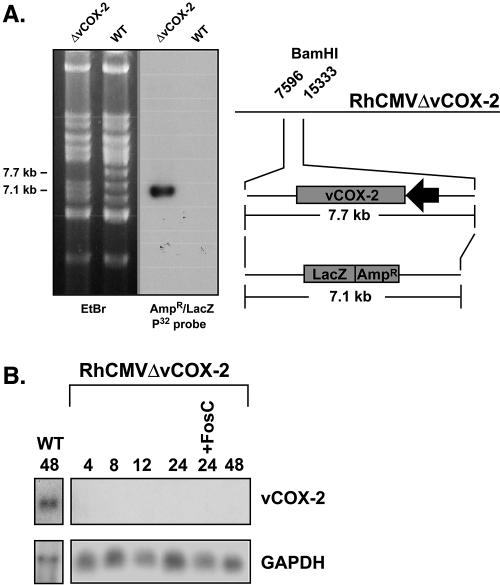
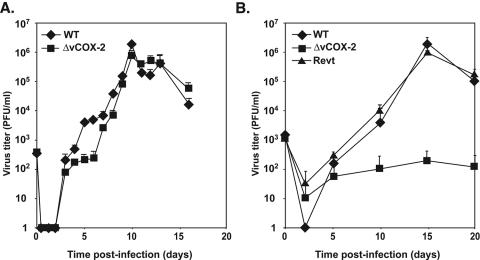
Cyclooxygenase-2 (COX-2) is a cellular enzyme in the eicosanoid synthetic pathway that mediates the synthesis of prostaglandins from arachidonic acid. The eicosanoids function as critical regulators of a number of cellular processes, including the acute and chronic inflammatory response, hemostasis, and the innate immune response. Human cytomegalovirus (HCMV), which does not encode a viral COX-2 isoform, has been shown to induce cellular COX-2 expression. Importantly, although the precise role of COX-2 in CMV replication is unknown, COX-2 induction was shown to be critical for normal HCMV replication. In an earlier study, we identified an open reading frame (Rh10) within the rhesus cytomegalovirus (RhCMV) genome that encoded a putative protein (designated vCOX-2) with high homology to cellular COX-2. In the current study, we show that vCOX-2 is expressed with early-gene kinetics during RhCMV infection, resulting in production of a 70-kDa protein. Consistent with the expression of a viral COX-2 isoform, cellular COX-2 expression was not induced during RhCMV infection. Finally, analysis of growth of recombinant RhCMV with vCOX-2 deleted identified vCOX-2 as a critical determinant for replication in endothelial cells.
Figures
Similar articles
-
Limited dissemination and shedding of the UL128 complex-intact, UL/b'-defective rhesus cytomegalovirus strain 180.92.J Virol. 2014 Aug;88(16):9310-20. doi: 10.1128/JVI.00162-14. Epub 2014 Jun 4. J Virol. 2014. PMID: 24899204 Free PMC article.
-
Inactivation of the Human Cytomegalovirus US20 Gene Hampers Productive Viral Replication in Endothelial Cells.J Virol. 2015 Nov;89(21):11092-106. doi: 10.1128/JVI.01141-15. Epub 2015 Aug 26. J Virol. 2015. PMID: 26311874 Free PMC article.
-
Complete sequence and genomic analysis of rhesus cytomegalovirus.J Virol. 2003 Jun;77(12):6620-36. doi: 10.1128/jvi.77.12.6620-6636.2003. J Virol. 2003. PMID: 12767982 Free PMC article.
-
Many actions of cyclooxygenase-2 in cellular dynamics and in cancer.J Cell Physiol. 2002 Mar;190(3):279-86. doi: 10.1002/jcp.10068. J Cell Physiol. 2002. PMID: 11857443 Review.
-
The role of cyclooxygenase-2 in the pathogenesis of skin cancer.J Drugs Dermatol. 2002 Jul;1(1):44-7. J Drugs Dermatol. 2002. PMID: 12847753 Review.
Cited by
-
A novel strain of cynomolgus macaque cytomegalovirus: implications for host-virus co-evolution.BMC Genomics. 2016 Apr 5;17:277. doi: 10.1186/s12864-016-2588-3. BMC Genomics. 2016. PMID: 27044312 Free PMC article.
-
Inhibition of cyclooxygenase activity blocks cell-to-cell spread of human cytomegalovirus.Proc Natl Acad Sci U S A. 2008 Dec 9;105(49):19468-73. doi: 10.1073/pnas.0810740105. Epub 2008 Nov 25. Proc Natl Acad Sci U S A. 2008. PMID: 19033472 Free PMC article.
-
Meal for Two: Human Cytomegalovirus-Induced Activation of Cellular Metabolism.Viruses. 2019 Mar 19;11(3):273. doi: 10.3390/v11030273. Viruses. 2019. PMID: 30893762 Free PMC article. Review.
-
Efficient replication of rhesus cytomegalovirus variants in multiple rhesus and human cell types.Proc Natl Acad Sci U S A. 2008 Dec 16;105(50):19950-5. doi: 10.1073/pnas.0811063106. Epub 2008 Dec 8. Proc Natl Acad Sci U S A. 2008. PMID: 19064925 Free PMC article.
-
Macaque models of human infectious disease.ILAR J. 2008;49(2):220-55. doi: 10.1093/ilar.49.2.220. ILAR J. 2008. PMID: 18323583 Free PMC article. Review.
References
-
- Brune, W., C. Menard, J. Heesemann, and U. H. Koszinowski. 2001. A ribonucleotide reductase homolog of cytomegalovirus and endothelial cell tropism. Science 291:303-305. - PubMed
-
- Bubeck, A., U. Reusch, M. Wagner, T. Ruppert, W. Muranyi, P. M. Kloetzel, and U. H. Koszinowski. 2002. The glycoprotein gp48 of murine cytomegalovirus L proteasome-dependent cytosolic dislocation and degradation. J. Biol. Chem. 277:2216-2224. - PubMed
-
- Burleigh, M. E., V. R. Babaev, J. A. Oates, R. C. Harris, S. Gautam, D. Riendeau, L. J. Marnett, J. D. Morrow, S. Fazio, and M. F. Linton. 2002. Cyclooxygenase-2 promotes early atherosclerotic lesion formation in LDL receptor-deficient mice. Circulation 105:1816-1823. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
