

Conditionally replicating adenoviruses kill tumor cells via a basic apoptotic machinery-independent mechanism that resembles necrosis-like programmed cell death
- PMID: 15507611
- PMCID: PMC525077
- DOI: 10.1128/JVI.78.22.12243-12251.2004
Conditionally replicating adenoviruses kill tumor cells via a basic apoptotic machinery-independent mechanism that resembles necrosis-like programmed cell death
Abstract
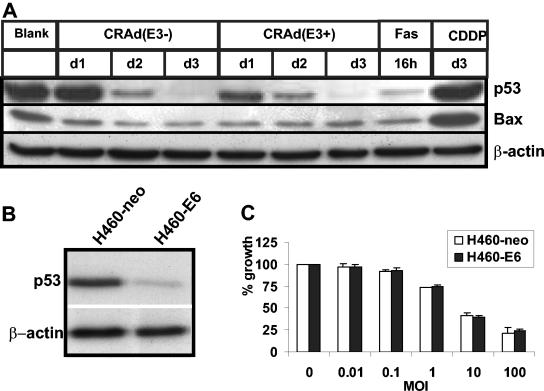
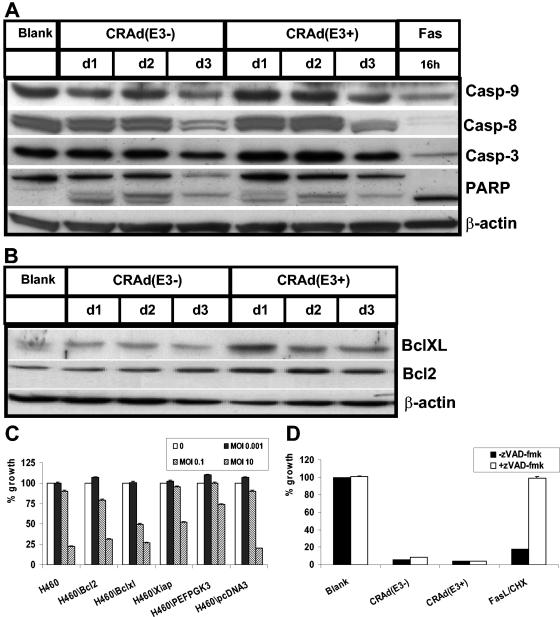
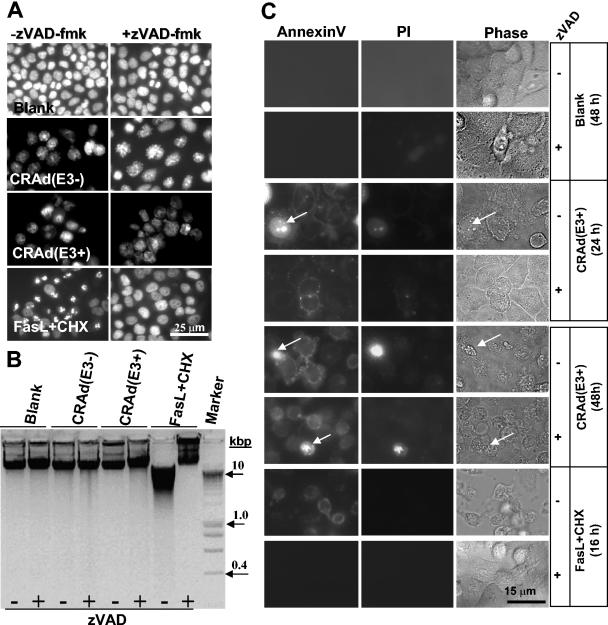
Conditionally replicating adenoviruses (CRAds) represent a promising class of novel anticancer agents that are used for virotherapy. The E1ADelta24 mutation-based viruses, Ad5-Delta24 [CRAd(E3-); E3 region deleted] and infectivity-enhanced Ad5-Delta24RGD [CRAd(E3+)] have been shown to potently eradicate tumor cells. The presence of the E3 region in the latter virus is known to improve cell killing that can be attributed to the presence of the oncolysis-enhancing Ad death protein. The more precise mechanism by which CRAds kill tumor cells is unclear, and the role of the host cell apoptotic machinery in this process has been addressed only in a limited way. Here, we examine the role of several major apoptotic pathways in the CRAd-induced killing of non-small-cell lung cancer H460 cells. As expected, CRAd(E3+) was more potent than CRAd(E3-). No evidence for the involvement of the p53-Bax apoptotic pathway was found. Western blot analyses demonstrated strong suppression of p53 expression and unchanged Bax levels during viral replication, and stable overexpression of human papillomavirus type 16-E6 in H460 cells did not affect killing by both CRAds. CRAd activity was also not hampered by stable overexpression of anti-apoptotic Bcl2 or BclXL, and endogenous Bcl2/BclXL protein levels remained constant during the oncolytic cycle. Some evidence for caspase processing was obtained at late time points after infection; however, the inhibition of caspases by the X-linked inhibitor of apoptosis protein overexpression or cotreatment with zVAD-fmk did not inhibit CRAd-dependent cell death. Analyses of several apoptotic features revealed no evidence for nuclear fragmentation or DNA laddering, although phosphatidylserine externalization was detected. We conclude that despite the known apoptosis-modulating abilities of individual Ad proteins, Ad5-Delta24-based CRAds trigger necrosis-like cell death. In addition, we propose that deregulated apoptosis in cancer cells, a possible drug resistance mechanism, provides no barrier for CRAd efficacy.
Figures

Similar articles
-
Conditionally replicative adenovirus expressing p53 exhibits enhanced oncolytic potency.Cancer Res. 2002 Nov 1;62(21):6165-71. Cancer Res. 2002. PMID: 12414643
-
Gene-directed enzyme prodrug therapy with carboxylesterase enhances the anticancer efficacy of the conditionally replicating adenovirus AdDelta24.Gene Ther. 2005 Jun;12(12):1011-8. doi: 10.1038/sj.gt.3302492. Gene Ther. 2005. PMID: 15729367
-
Inter-patient variation in efficacy of five oncolytic adenovirus candidates for ovarian cancer therapy.J Gene Med. 2004 Dec;6(12):1333-42. doi: 10.1002/jgm.635. J Gene Med. 2004. PMID: 15493039
-
Conditionally replicating adenoviruses as anticancer agents and ways to improve their efficacy.J Exp Ther Oncol. 2004 Apr;4(1):37-57. J Exp Ther Oncol. 2004. PMID: 15255291 Review.
-
Toward a new generation of conditionally replicating adenoviruses: pairing tumor selectivity with maximal oncolysis.Hum Gene Ther. 2002 Mar 1;13(4):485-95. doi: 10.1089/10430340252809784. Hum Gene Ther. 2002. PMID: 11874627 Review.
Cited by
-
Dual E1A oncolytic adenovirus: targeting tumor heterogeneity with two independent cancer-specific promoter elements, DF3/MUC1 and hTERT.Cancer Gene Ther. 2011 Mar;18(3):153-66. doi: 10.1038/cgt.2010.52. Epub 2010 Sep 24. Cancer Gene Ther. 2011. PMID: 20865021 Free PMC article.
-
Physical interventions to interrupt or reduce the spread of respiratory viruses.Cochrane Database Syst Rev. 2023 Jan 30;1(1):CD006207. doi: 10.1002/14651858.CD006207.pub6. Cochrane Database Syst Rev. 2023. PMID: 36715243 Free PMC article. Review.
-
Human adenovirus type 5 induces cell lysis through autophagy and autophagy-triggered caspase activity.J Virol. 2011 May;85(10):4720-9. doi: 10.1128/JVI.02032-10. Epub 2011 Mar 2. J Virol. 2011. PMID: 21367888 Free PMC article.
-
Silencing of Mcl-1 overcomes resistance of melanoma cells against TRAIL-armed oncolytic adenovirus by enhancement of apoptosis.J Mol Med (Berl). 2021 Sep;99(9):1279-1291. doi: 10.1007/s00109-021-02081-3. Epub 2021 May 24. J Mol Med (Berl). 2021. PMID: 34028599 Free PMC article.
-
Oncolytic viruses in cancer therapy.Cancer Lett. 2007 Sep 8;254(2):178-216. doi: 10.1016/j.canlet.2007.02.002. Epub 2007 Mar 23. Cancer Lett. 2007. PMID: 17383089 Free PMC article. Review.
References
-
- Adams, J. M., and S. Cory. 2001. Life-or-death decisions by the Bcl-2 protein family. Trends Biochem. Sci. 26:61-66. - PubMed
-
- Alemany, R., C. Balague, and D. T. Curiel. 2000. Replicative adenoviruses for cancer therapy. Nat. Biotechnol. 18:723-727. - PubMed
-
- Bauerschmitz, G. J., J. T. Lam, A. Kanerva, K. Suzuki, D. M. Nettelbeck, I. Dmitriev, V. Krasnykh, G. V. Mikheeva, M. N. Barnes, R. D. Alvarez, P. Dall, R. Alemany, D. T. Curiel, and A. Hemminki. 2002. Treatment ovarian cancer with a tropism modified oncolytic adenovirus. Cancer Res. 62:1266-1270. - PubMed
-
- Boulakia, C. A., G. Chen, F. W. Ng, J. G. Teodoro, P. E. Branton, D. W. Nicholson, G. G. Poirier, and G. C. Shore. 1996. Bcl-2 and adenovirus E1B 19 kDA protein prevent E1A-induced processing of CPP32 and cleavage of poly(ADP-ribose) polymerase. Oncogene 12:529-535. - PubMed
-
- Boyd, J. M., S. Malstrom, T. Subramanian, L. K. Venkatesh, U. Schaeper, B. Elangovan, C. D'Sa-Eipper, and G. Chinnadurai. 1994. Adenovirus E1B 19 kDa and Bcl-2 proteins interact with a common set of cellular proteins. Cell 79:341-351. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
