

Review
doi: 10.1016/j.pbiomolbio.2004.06.001.
Tonic excitation and inhibition of neurons: ambient transmitter sources and computational consequences
Affiliations
- PMID: 15471587
- PMCID: PMC8906495
- DOI: 10.1016/j.pbiomolbio.2004.06.001
Item in Clipboard
Review
Tonic excitation and inhibition of neurons: ambient transmitter sources and computational consequences
Prog Biophys Mol Biol.
2005 Jan.
Abstract
Tonic activation of excitatory and inhibitory receptors, by the ambient concentration of neurotransmitters in the extracellular space of the brain, has been suggested to underlie phenomena as diverse as relapse to cocaine use by reward pathways in the striatum, sparse coding of motor information in the cerebellum, and control of the development of the cerebral and cerebellar cortices. Here we assess the mechanisms which may determine the ambient levels of excitatory and inhibitory neurotransmitters, and consider their likely effect on information processing.
Figures
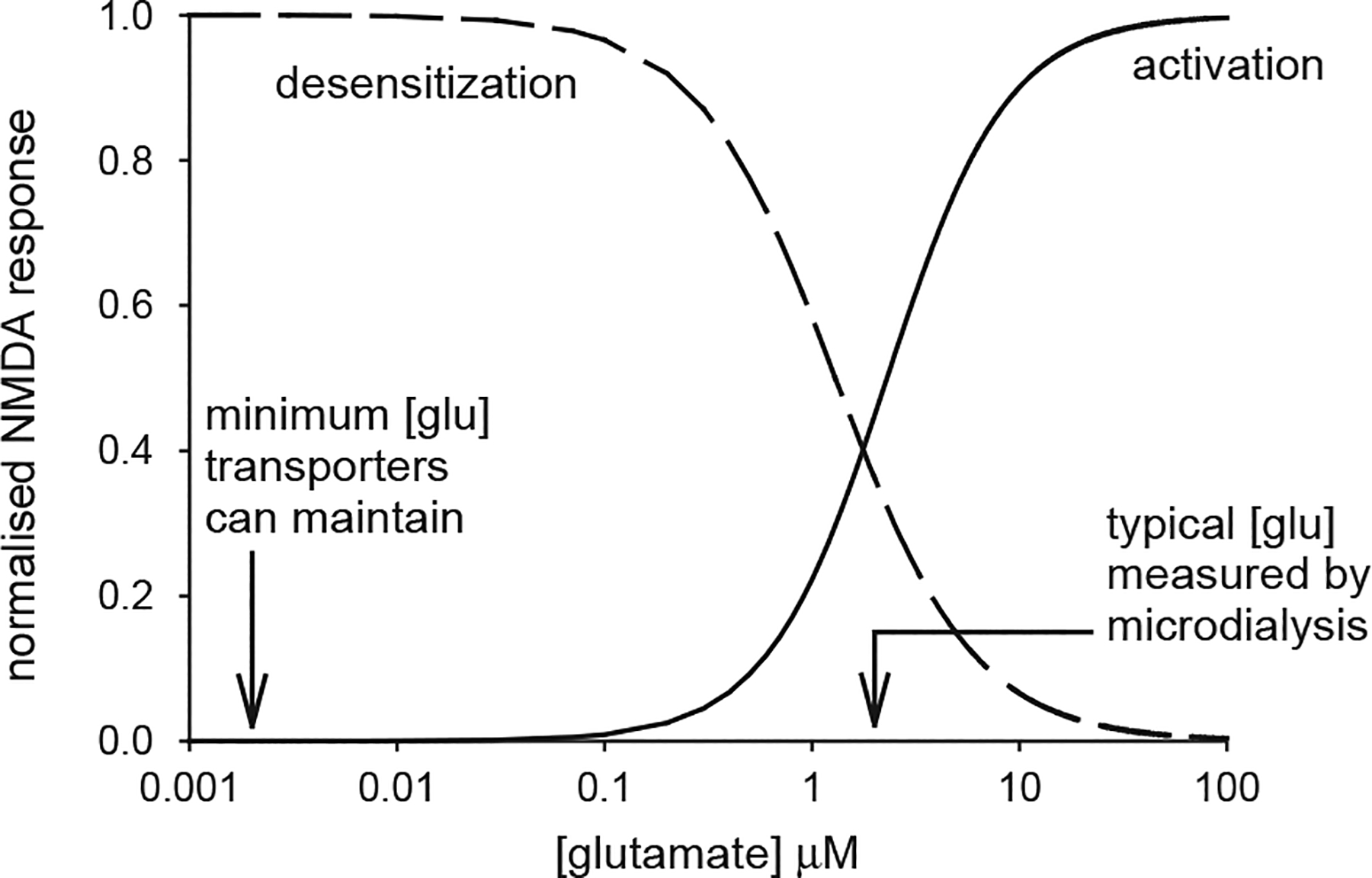
The effect of a non-zero baseline extracellular glutamate concentration on the activation of NMDA receptors. Continuous curve is the activation curve for NMDA receptors (, with EC50 = 2.3 μM, from Patneau and Mayer, 1990). Na+-dependent transporters with the stoichiometry measured for EAAC1 and GLT-1 can theoretically lower [glu]o to ~2 nM (left arrow). Microdialysis experiments typically measure a resting [glu]o of ~2 μM (right arrow), which is sufficient to activate NMDA receptors by 45% of their maximum activation, so that subsequent rises of [glu]o produce a smaller change of activation than would occur with no glutamate present initially. Prolonged presence of glutamate also desensitises NMDA receptors. The dashed line, given by , with IC50=1.3 μM, shows the inhibition of the NMDA component of EPSCs measured by Zorumski et al. (1996): this curve overestimates the amount of desensitization occurring, because pre-synaptic actions of glutamate contribute a small amount to the EPSC depression (e.g. at 1 μM glutamate, presynaptic inhibition alone inhibits the response by about 12% while the combination of pre-synaptic inhibition and desensitization inhibits by about 42%: see Zorumski et al. (1996) Fig. 6B).
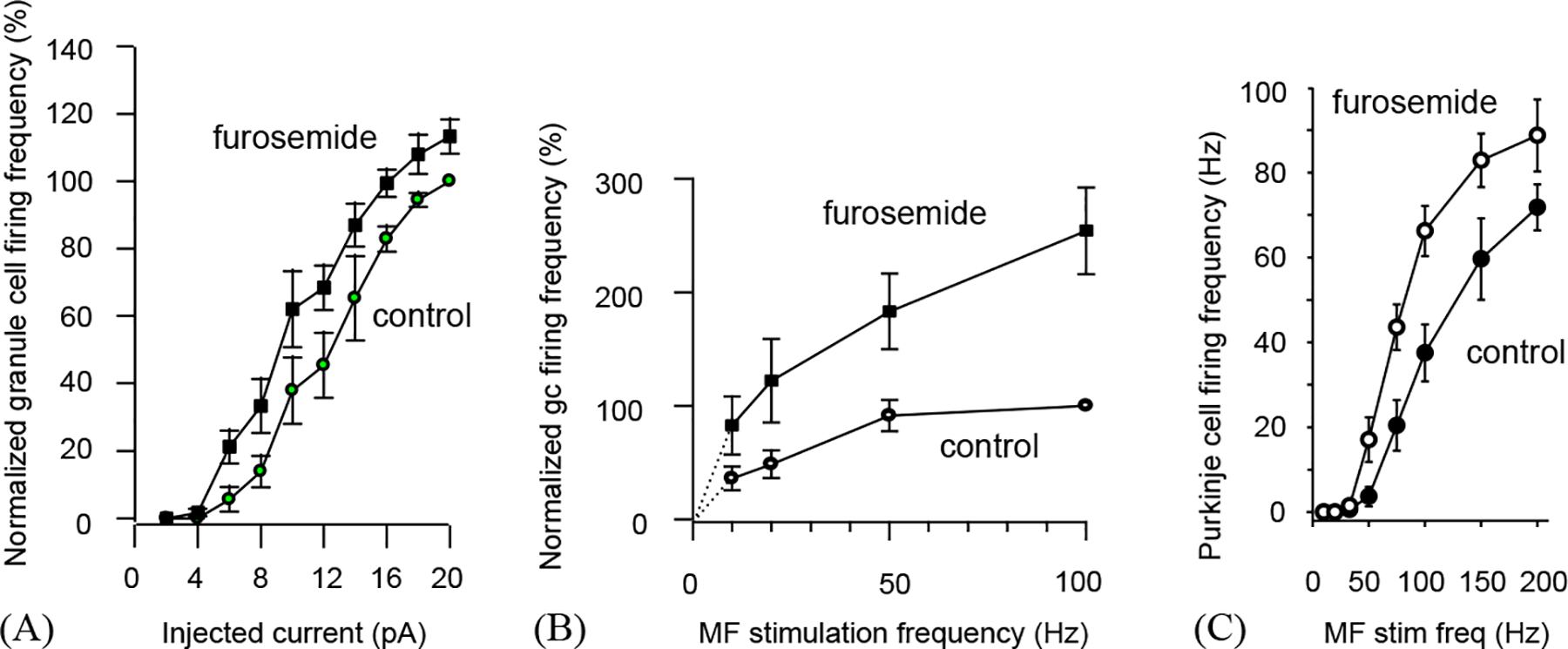
The effect of tonic inhibition of cerebellar granule cells on information transfer through the cerebellar cortex of rats (from the study of Hamann et al., 2002). (A) Granule cell firing rate (normalised to maximum rate in nine cells) as a function of injected current, in control conditions and with tonic inhibition by high affinity GABAA receptors blocked with furosemide. The input–output curve is shifted to the left by blocking the tonic inhibition. (B) Granule cell firing rate (normalized to maximum rate in four cells) as a function of mossy fibre stimulation rate, in the presence and absence (furosemide) of tonic inhibition. (C) Purkinje cell firing rate as a function of mossy fibre stimulation rate, in the presence and absence (furosemide) of tonic inhibition.
Similar articles
-
Update on the glutamatergic neurotransmitter system and the role of excitotoxicity in amyotrophic lateral sclerosis.Muscle Nerve. 2002 Oct;26(4):438-58. doi: 10.1002/mus.10186. Muscle Nerve. 2002. PMID: 12362409 Review.
-
Glutamate receptors in the central nervous system.Ann N Y Acad Sci. 1993 Dec 20;707:126-35. doi: 10.1111/j.1749-6632.1993.tb38048.x. Ann N Y Acad Sci. 1993. PMID: 9137548 Review. No abstract available.
-
Evidence that functional glutamate receptors are not expressed on rat or human cerebromicrovascular endothelial cells.J Cereb Blood Flow Metab. 1998 Apr;18(4):396-406. doi: 10.1097/00004647-199804000-00008. J Cereb Blood Flow Metab. 1998. PMID: 9538905
-
GABA-dependent firing of glutamate-evoked action potentials at AMPA/kainate receptors in developing hypothalamic neurons.J Neurophysiol. 1998 Feb;79(2):716-26. doi: 10.1152/jn.1998.79.2.716. J Neurophysiol. 1998. PMID: 9463435
-
Postsynaptic modulation of AMPA- and NMDA-receptor currents by Group III metabotropic glutamate receptors in rat nucleus accumbens.Brain Res. 2003 Jun 20;976(1):60-8. doi: 10.1016/s0006-8993(03)02676-3. Brain Res. 2003. PMID: 12763622
Cited by
-
Multiple and plastic receptors mediate tonic GABAA receptor currents in the hippocampus.J Neurosci. 2005 Oct 26;25(43):10016-24. doi: 10.1523/JNEUROSCI.2520-05.2005. J Neurosci. 2005. PMID: 16251450 Free PMC article.
-
Higher ambient synaptic glutamate at inhibitory versus excitatory neurons differentially impacts NMDA receptor activity.Nat Commun. 2018 Oct 1;9(1):4000. doi: 10.1038/s41467-018-06512-7. Nat Commun. 2018. PMID: 30275542 Free PMC article.
-
Tonic activation of NMDA receptors by ambient glutamate of non-synaptic origin in the rat hippocampus.J Physiol. 2007 Apr 15;580(Pt. 2):373-83. doi: 10.1113/jphysiol.2006.123570. Epub 2006 Dec 21. J Physiol. 2007. PMID: 17185337 Free PMC article.
-
Short and Long-Term Attentional Firing Rates Can Be Explained by ST-Neuron Dynamics.Front Neurosci. 2018 Mar 2;12:123. doi: 10.3389/fnins.2018.00123. eCollection 2018. Front Neurosci. 2018. PMID: 29551961 Free PMC article.
-
A Cytotoxic, Co-operative Interaction Between Energy Deprivation and Glutamate Release From System xc- Mediates Aglycemic Neuronal Cell Death.ASN Neuro. 2015 Nov 9;7(6):1759091415614301. doi: 10.1177/1759091415614301. Print 2015 Nov-Dec. ASN Neuro. 2015. PMID: 26553727 Free PMC article.
References
-
- Attwell D, Barbour B, Szatkowski M, 1993. Nonvesicular release of neurotransmitter. Neuron 11, 401–407. - PubMed
-
- Baker DA, McFarland K, Lake RW, Shen H, Tang XC, Toda S, Kalivas PW, 2003. Neuroadaptations in cystine-glutamate exchange underlie cocaine relapse. Nat. Neurosci 6, 743–749. - PubMed
-
- Banke TG, Traynelis SF, 2003. Activation of NR1/NR2B NMDA receptors. Nat. Neurosci 6, 144–152. - PubMed
-
- Bannai S, 1984. Transport of cystine and cysteine in mammalian cells. Biochem. Biophys. Acta 779, 289–306. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
