

Dual sensitivity of sarcoplasmic/endoplasmic Ca2+-ATPase to cytosolic and endoplasmic reticulum Ca2+ as a mechanism of modulating cytosolic Ca2+ oscillations
- PMID: 15260801
- PMCID: PMC1134077
- DOI: 10.1042/BJ20040629
Dual sensitivity of sarcoplasmic/endoplasmic Ca2+-ATPase to cytosolic and endoplasmic reticulum Ca2+ as a mechanism of modulating cytosolic Ca2+ oscillations
Abstract
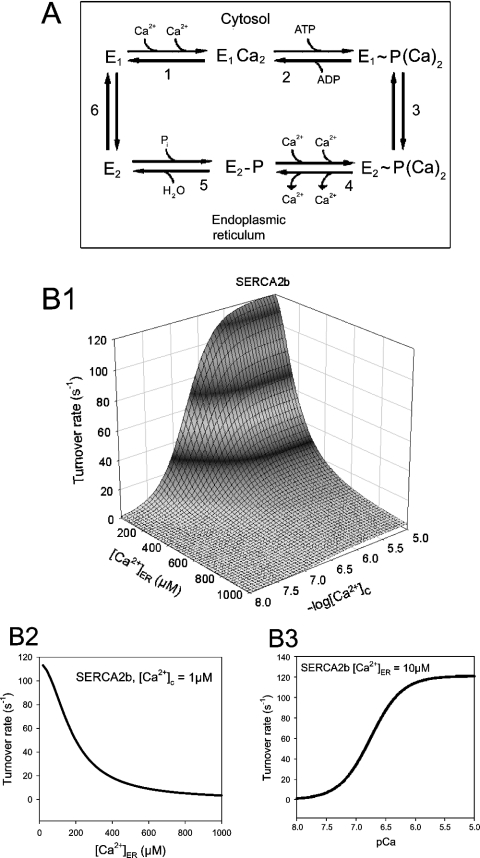
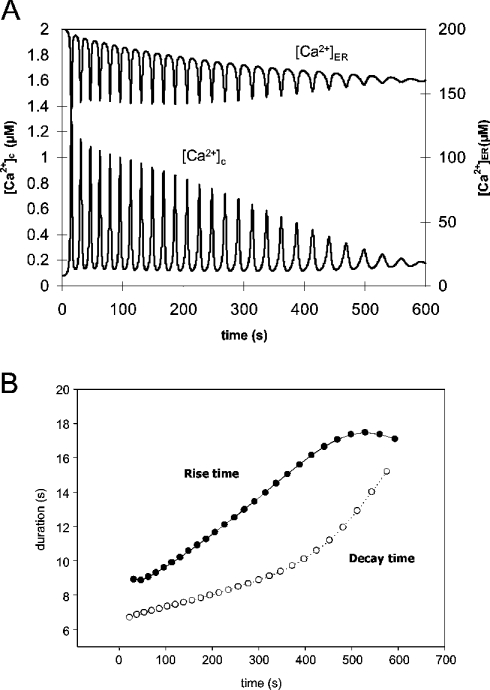
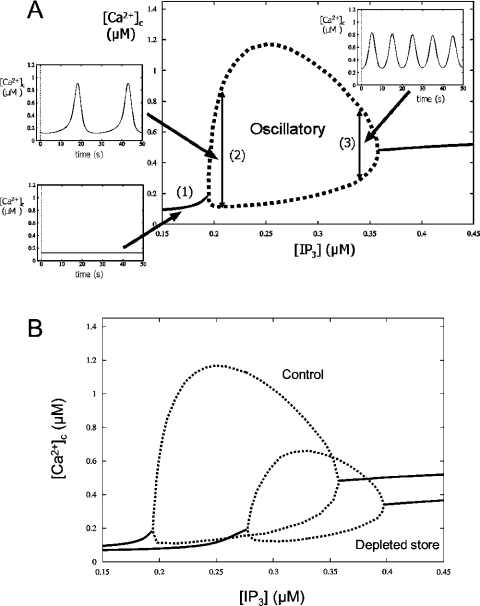
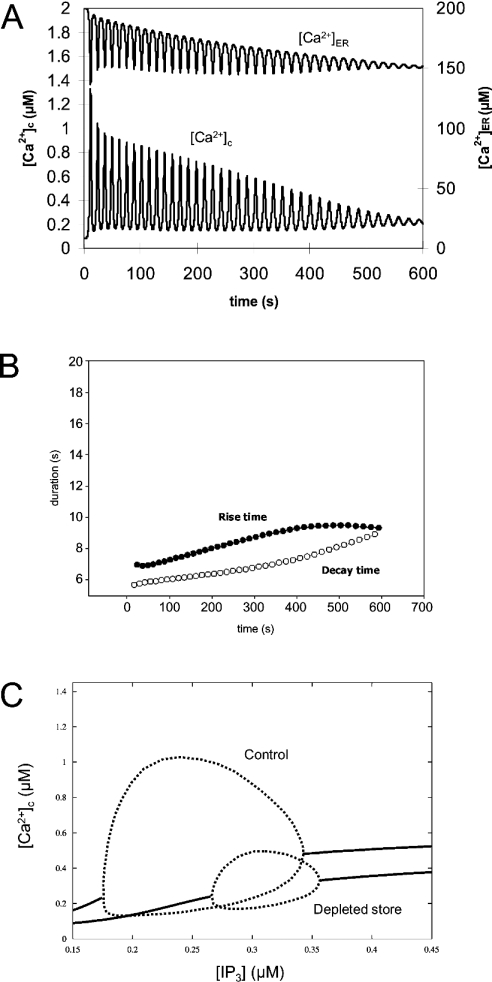
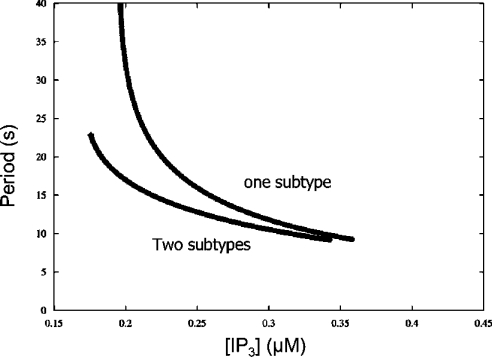
The effects of ER (endoplasmic reticulum) Ca2+ on cytosolic Ca2+ oscillations in pancreatic acinar cells were investigated using mathematical models of the Ca2+ oscillations. We first examined the mathematical model of SERCA (sarcoplasmic/endoplasmic reticulum Ca2+-ATPase) to reproduce the highly co-operative inhibitory effect of Ca2+ in the ER lumen on ER Ca2+ uptake in the acinar cells. The model predicts that luminal Ca2+ would most probably inhibit the conversion of the conformation state with luminal Ca2+-binding sites (E2) into the conformation state with cytoplasmic Ca2+-binding sites (E1). The SERCA model derived from this prediction showed dose-response relationships to cytosolic and luminal Ca2+ concentrations that were consistent with the experimental data from the acinar cells. According to a mathematical model of cytosolic Ca2+ oscillations based on the modified SERCA model, a small decrease in the concentration of endoplasmic reticulum Ca2+ (approx. 20% of the total) was sufficient to abolish the oscillations. When a single type of IP3R (IP3 receptor) was included in the model, store depletion decreased the spike frequency. However, the frequency became less sensitive to store depletion when we added another type of IP3R with higher sensitivity to the concentration of free Ca2+ in the cytosol. Bifurcation analysis of the mathematical model showed that the loss of Ca2+ from the ER lumen decreased the sensitivity of cytosolic Ca2+ oscillations to IP3 [Ins(1,4,5)P3]. The addition of a high-affinity IP3R did not alter this property, but significantly decreased the sensitivity of the spike frequency to IP3. Our mathematical model demonstrates how luminal Ca2+, through its effect on Ca2+ uptake, can control cytosolic Ca2+ oscillations.
Figures
Similar articles
-
Modeling the impact of store-operated Ca2+ entry on intracellular Ca2+ oscillations.Math Biosci. 2006 Dec;204(2):232-49. doi: 10.1016/j.mbs.2006.03.001. Epub 2006 Apr 19. Math Biosci. 2006. PMID: 16620876
-
On the roles of Ca2+ diffusion, Ca2+ buffers, and the endoplasmic reticulum in IP3-induced Ca2+ waves.Biophys J. 1995 Nov;69(5):2139-53. doi: 10.1016/S0006-3495(95)80088-3. Biophys J. 1995. PMID: 8580358 Free PMC article.
-
Organization of Ca2+ stores in myeloid cells: association of SERCA2b and the type-1 inositol-1,4,5-trisphosphate receptor.Biochem J. 1996 May 15;316 ( Pt 1)(Pt 1):137-42. doi: 10.1042/bj3160137. Biochem J. 1996. PMID: 8645196 Free PMC article.
-
Sarco-Endoplasmic Reticulum Calcium Release Model Based on Changes in the Luminal Calcium Content.Adv Exp Med Biol. 2020;1131:337-370. doi: 10.1007/978-3-030-12457-1_14. Adv Exp Med Biol. 2020. PMID: 31646517 Review.
-
Calcium transport pathways of pancreatic acinar cells.Annu Rev Physiol. 1989;51:83-105. doi: 10.1146/annurev.ph.51.030189.000503. Annu Rev Physiol. 1989. PMID: 2540704 Review.
Cited by
-
Modulation of endoplasmic reticulum Ca2+ store filling by cyclic ADP-ribose promotes inositol trisphosphate (IP3)-evoked Ca2+ signals.J Biol Chem. 2010 Aug 6;285(32):25053-61. doi: 10.1074/jbc.M109.095257. Epub 2010 Jun 10. J Biol Chem. 2010. PMID: 20538594 Free PMC article.
-
Mechanical and biochemical modeling of cortical oscillations in spreading cells.Biophys J. 2008 Jun;94(12):4605-20. doi: 10.1529/biophysj.107.121335. Epub 2008 Mar 7. Biophys J. 2008. PMID: 18326667 Free PMC article.
-
Phe27Cys polymorphism of the human delta opioid receptor predisposes cells to compromised calcium signaling.Mol Cell Biochem. 2011 May;351(1-2):173-81. doi: 10.1007/s11010-011-0725-5. Epub 2011 Jan 14. Mol Cell Biochem. 2011. PMID: 21234650
-
A buffering SERCA pump in models of calcium dynamics.Biophys J. 2006 Jul 1;91(1):151-63. doi: 10.1529/biophysj.105.075747. Epub 2006 Apr 14. Biophys J. 2006. PMID: 16617079 Free PMC article.
-
Altered Endoplasmic Reticulum Calcium Pump Expression during Breast Tumorigenesis.Breast Cancer (Auckl). 2011;5:163-74. doi: 10.4137/BCBCR.S7481. Epub 2011 Jul 14. Breast Cancer (Auckl). 2011. PMID: 21863130 Free PMC article.
References
-
- Tepikin A. V., Voronina S. G., Gallacher D. V., Petersen O. H. Pulsatile Ca2+ extrusion from single pancreatic acinar cells during receptor-activated cytosolic Ca2+ spiking. J. Biol. Chem. 1992;267:14073–14076. - PubMed
-
- Favre C. J., Schrenzel J., Jacquet J., Lew D. P., Krause K. H. Highly supralinear feedback inhibition of Ca2+ uptake by the Ca2+ load of intracellular stores. J. Biol. Chem. 1996;271:14925–14930. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous