

Drosophila nipped-B protein supports sister chromatid cohesion and opposes the stromalin/Scc3 cohesion factor to facilitate long-range activation of the cut gene
- PMID: 15060134
- PMCID: PMC381657
- DOI: 10.1128/MCB.24.8.3100-3111.2004
Drosophila nipped-B protein supports sister chromatid cohesion and opposes the stromalin/Scc3 cohesion factor to facilitate long-range activation of the cut gene
Abstract
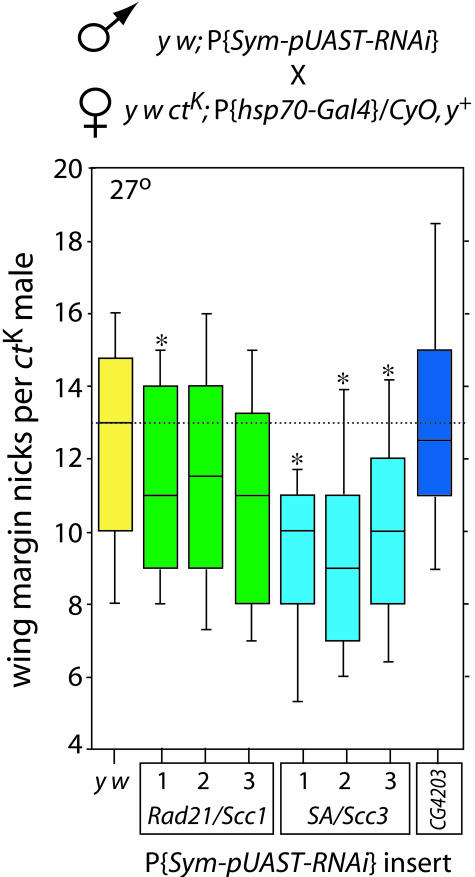
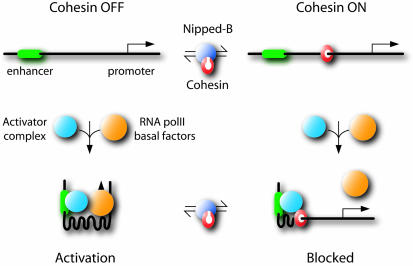
The Drosophila melanogaster Nipped-B protein facilitates transcriptional activation of the cut and Ultrabithorax genes by remote enhancers. Sequence homologues of Nipped-B, Scc2 of Saccharomyces cerevisiae, and Mis4 of Schizosaccharomyces pombe are required for sister chromatid cohesion during mitosis. The evolutionarily conserved Cohesin protein complex mediates sister chromatid cohesion, and Scc2 and Mis4 are needed for Cohesin to associate with chromosomes. Here, we show that Nipped-B is also required for sister chromatid cohesion but that, opposite to the effect of Nipped-B, the stromalin/Scc3 component of Cohesin inhibits long-range activation of cut. To explain these findings, we propose a model based on the chromatin domain boundary activities of Cohesin in which Nipped-B facilitates cut activation by alleviating Cohesin-mediated blocking of enhancer-promoter communication.
Figures
Similar articles
-
Functional links between Drosophila Nipped-B and cohesin in somatic and meiotic cells.Chromosoma. 2008 Feb;117(1):51-66. doi: 10.1007/s00412-007-0125-5. Epub 2007 Oct 2. Chromosoma. 2008. PMID: 17909832 Free PMC article.
-
Effects of sister chromatid cohesion proteins on cut gene expression during wing development in Drosophila.Development. 2005 Nov;132(21):4743-53. doi: 10.1242/dev.02064. Epub 2005 Oct 5. Development. 2005. PMID: 16207752 Free PMC article.
-
Nipped-B, a Drosophila homologue of chromosomal adherins, participates in activation by remote enhancers in the cut and Ultrabithorax genes.Genetics. 1999 Jun;152(2):577-93. doi: 10.1093/genetics/152.2.577. Genetics. 1999. PMID: 10353901 Free PMC article.
-
Cohesin, gene expression and development: lessons from Drosophila.Chromosome Res. 2009;17(2):185-200. doi: 10.1007/s10577-009-9022-5. Chromosome Res. 2009. PMID: 19308700 Free PMC article. Review.
-
The Many Roles of Cohesin in Drosophila Gene Transcription.Trends Genet. 2019 Jul;35(7):542-551. doi: 10.1016/j.tig.2019.04.002. Epub 2019 May 23. Trends Genet. 2019. PMID: 31130395 Free PMC article. Review.
Cited by
-
PCNA promotes context-specific sister chromatid cohesion establishment separate from that of chromatin condensation.Cell Cycle. 2020 Oct;19(19):2436-2450. doi: 10.1080/15384101.2020.1804221. Epub 2020 Sep 14. Cell Cycle. 2020. PMID: 32926661 Free PMC article.
-
NIPBL mutational analysis in 120 individuals with Cornelia de Lange syndrome and evaluation of genotype-phenotype correlations.Am J Hum Genet. 2004 Oct;75(4):610-23. doi: 10.1086/424698. Epub 2004 Aug 18. Am J Hum Genet. 2004. PMID: 15318302 Free PMC article.
-
Multiple organ system defects and transcriptional dysregulation in the Nipbl(+/-) mouse, a model of Cornelia de Lange Syndrome.PLoS Genet. 2009 Sep;5(9):e1000650. doi: 10.1371/journal.pgen.1000650. Epub 2009 Sep 18. PLoS Genet. 2009. PMID: 19763162 Free PMC article.
-
Coming full circle: On the origin and evolution of the looping model for enhancer-promoter communication.J Biol Chem. 2022 Aug;298(8):102117. doi: 10.1016/j.jbc.2022.102117. Epub 2022 Jun 9. J Biol Chem. 2022. PMID: 35691341 Free PMC article. Review.
-
Cornelia de Lange Syndrome mutations in SMC1A cause cohesion defects in yeast.Genetics. 2023 Oct 4;225(2):iyad159. doi: 10.1093/genetics/iyad159. Genetics. 2023. PMID: 37650609 Free PMC article.
References
-
- Adams, M. D., et al. 2000. The genome sequence of Drosophila melanogaster. Science 287:2185-2195. - PubMed
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. - PubMed
-
- Blat, Y., and N. Kleckner. 1999. Cohesins bind to preferential sites along yeast chromosome III, with differential regulation along arms versus the centric region. Cell 98:249-259. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous