

S4 movement in a mammalian HCN channel
- PMID: 14676284
- PMCID: PMC2217412
- DOI: 10.1085/jgp.200308916
S4 movement in a mammalian HCN channel
Abstract
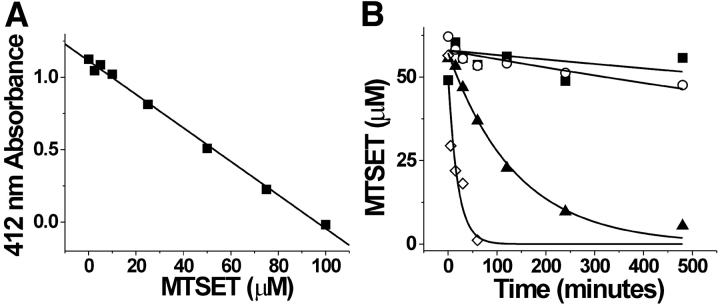
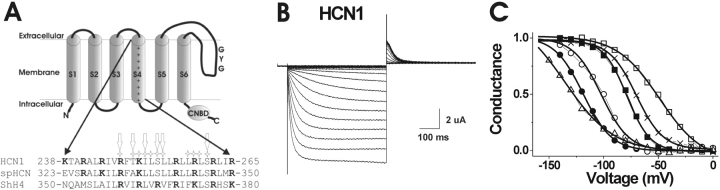
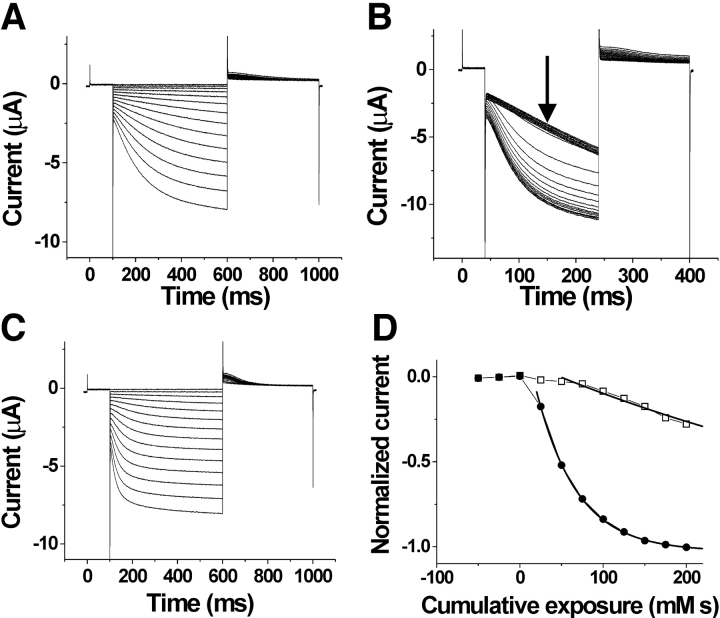
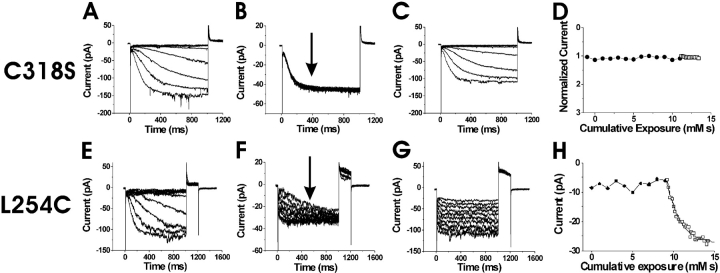
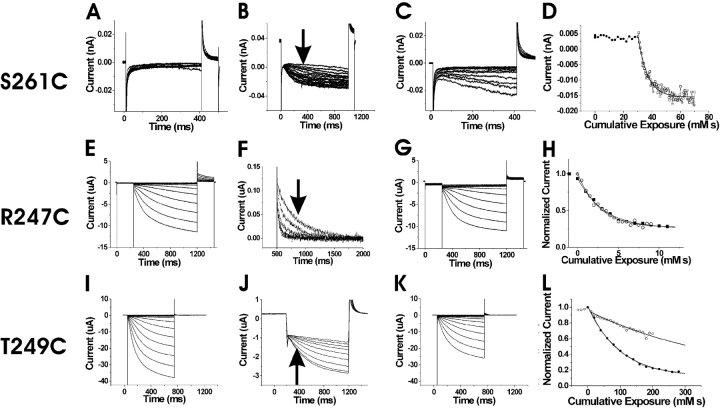
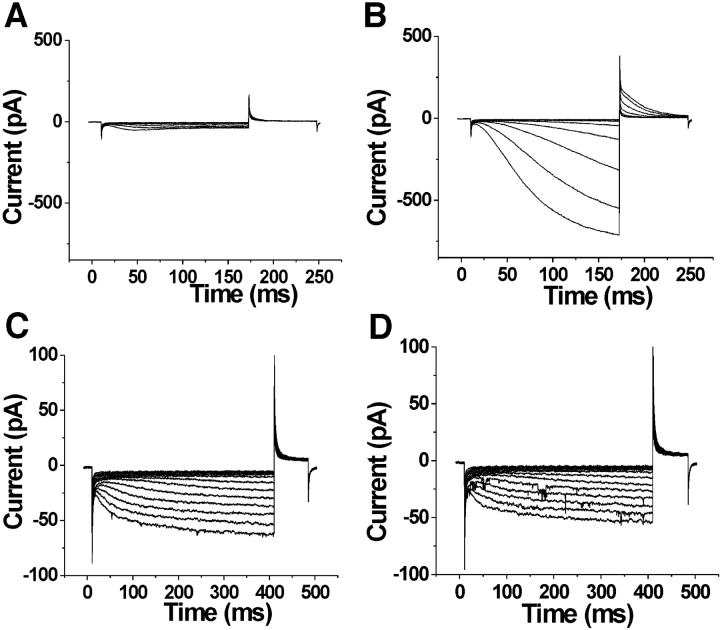
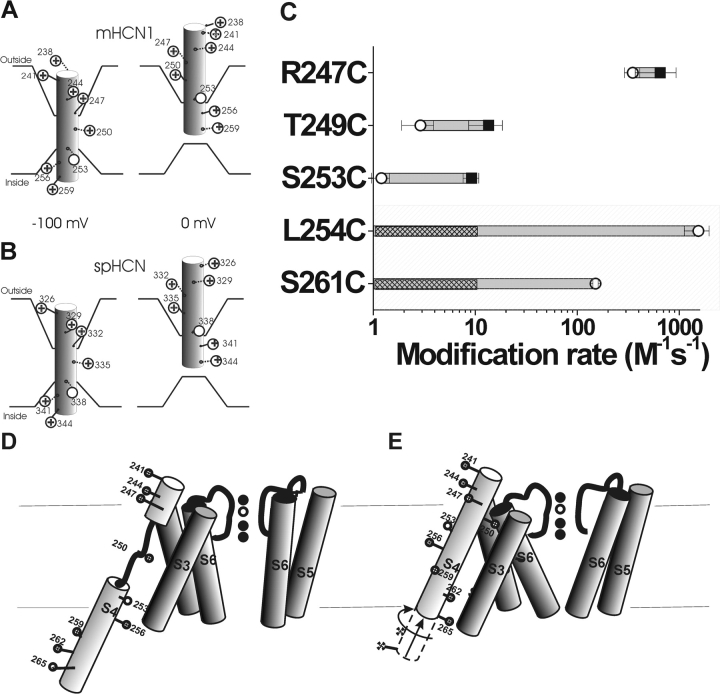
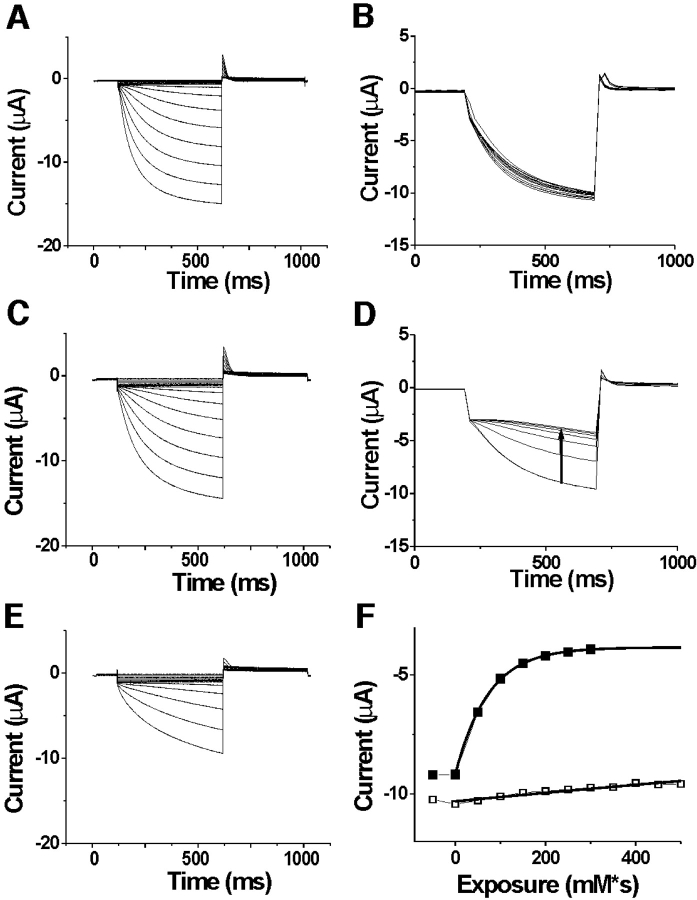
Hyperpolarization-activated, cyclic nucleotide-gated ion channels (HCN) mediate an inward cation current that contributes to spontaneous rhythmic firing activity in the heart and the brain. HCN channels share sequence homology with depolarization-activated Kv channels, including six transmembrane domains and a positively charged S4 segment. S4 has been shown to function as the voltage sensor and to undergo a voltage-dependent movement in the Shaker K+ channel (a Kv channel) and in the spHCN channel (an HCN channel from sea urchin). However, it is still unknown whether S4 undergoes a similar movement in mammalian HCN channels. In this study, we used cysteine accessibility to determine whether there is voltage-dependent S4 movement in a mammalian HCN1 channel. Six cysteine mutations (R247C, T249C, I251C, S253C, L254C, and S261C) were used to assess S4 movement of the heterologously expressed HCN1 channel in Xenopus oocytes. We found a state-dependent accessibility for four S4 residues: T249C and S253C from the extracellular solution, and L254C and S261C from the internal solution. We conclude that S4 moves in a voltage-dependent manner in HCN1 channels, similar to its movement in the spHCN channel. This S4 movement suggests that the role of S4 as a voltage sensor is conserved in HCN channels. In addition, to determine the reason for the different cAMP modulation and the different voltage range of activation in spHCN channels compared with HCN1 channels, we constructed a COOH-terminal-deleted spHCN. This channel appeared to be similar to a COOH-terminal-deleted HCN1 channel, suggesting that the main functional differences between spHCN and HCN1 channels are due to differences in their COOH termini or in the interaction between the COOH terminus and the rest of the channel protein in spHCN channels compared with HCN1 channels.
Figures
Similar articles
-
Changes in local S4 environment provide a voltage-sensing mechanism for mammalian hyperpolarization-activated HCN channels.J Gen Physiol. 2004 Jan;123(1):5-19. doi: 10.1085/jgp.200308918. Epub 2003 Dec 15. J Gen Physiol. 2004. PMID: 14676285 Free PMC article.
-
Voltage-sensing mechanism is conserved among ion channels gated by opposite voltages.Nature. 2002 Oct 24;419(6909):837-41. doi: 10.1038/nature01038. Nature. 2002. PMID: 12397358
-
Slow conformational changes of the voltage sensor during the mode shift in hyperpolarization-activated cyclic-nucleotide-gated channels.J Neurosci. 2007 Jan 10;27(2):270-8. doi: 10.1523/JNEUROSCI.3801-06.2007. J Neurosci. 2007. PMID: 17215386 Free PMC article.
-
Voltage sensing and activation gating of HCN pacemaker channels.Trends Cardiovasc Med. 2002 Jan;12(1):42-5. doi: 10.1016/s1050-1738(01)00141-4. Trends Cardiovasc Med. 2002. PMID: 11796244 Review.
-
Involvement of the S4-S5 linker and the C-linker domain regions to voltage-gating in plant Shaker channels: comparison with animal HCN and Kv channels.Plant Signal Behav. 2014;9(10):e972892. doi: 10.4161/15592316.2014.972892. Plant Signal Behav. 2014. PMID: 25482770 Free PMC article. Review.
Cited by
-
HCN Channel C-Terminal Region Speeds Activation Rates Independently of Autoinhibition.J Membr Biol. 2015 Dec;248(6):1043-60. doi: 10.1007/s00232-015-9816-7. Epub 2015 Jun 30. J Membr Biol. 2015. PMID: 26123597
-
Kinetic relationship between the voltage sensor and the activation gate in spHCN channels.J Gen Physiol. 2007 Jul;130(1):71-81. doi: 10.1085/jgp.200709769. J Gen Physiol. 2007. PMID: 17591986 Free PMC article.
-
Similar voltage-sensor movement in spHCN channels can cause closing, opening, or inactivation.J Gen Physiol. 2023 May 1;155(5):e202213170. doi: 10.1085/jgp.202213170. Epub 2023 Feb 8. J Gen Physiol. 2023. PMID: 36752823 Free PMC article.
-
A conformation change in the extracellular domain that accompanies desensitization of acid-sensing ion channel (ASIC) 3.J Gen Physiol. 2007 Apr;129(4):345-50. doi: 10.1085/jgp.200709757. J Gen Physiol. 2007. PMID: 17389250 Free PMC article.
-
Changes in local S4 environment provide a voltage-sensing mechanism for mammalian hyperpolarization-activated HCN channels.J Gen Physiol. 2004 Jan;123(1):5-19. doi: 10.1085/jgp.200308918. Epub 2003 Dec 15. J Gen Physiol. 2004. PMID: 14676285 Free PMC article.
References
-
- Aggarwal, S.K., and R. MacKinnon. 1996. Contribution of the S4 segment to gating charge in the Shaker K+ channel. Neuron. 16:1169–1177. - PubMed
-
- Baker, O.S., H.P. Larsson, L.M. Mannuzzu, and E.Y. Isacoff. 1998. Three transmembrane conformations and sequence-dependent displacement of the S4 domain in shaker K+ channel gating. Neuron. 20:1283–1294. - PubMed
-
- Chen, J., J.S. Mitcheson, M. Lin, and M.C. Sanguinetti. 2000. Functional roles of charged residues in the putative voltage sensor of the HCN2 pacemaker channel. J. Biol. Chem. 275:36465–36471. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
