

Telomere attachment, meiotic chromosome condensation, pairing, and bouquet stage duration are modified in spermatocytes lacking axial elements
- PMID: 14657244
- PMCID: PMC329396
- DOI: 10.1091/mbc.e03-07-0524
Telomere attachment, meiotic chromosome condensation, pairing, and bouquet stage duration are modified in spermatocytes lacking axial elements
Abstract
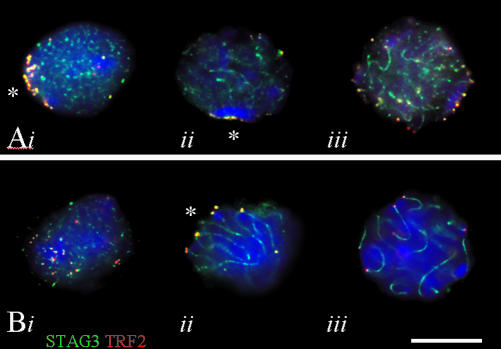
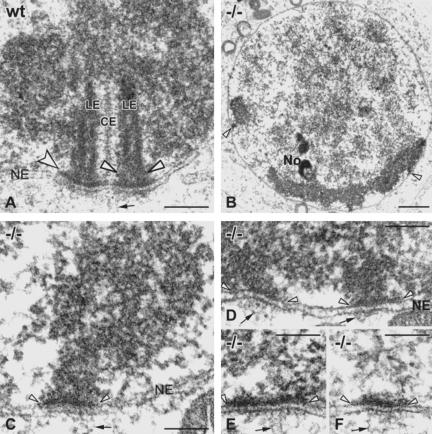
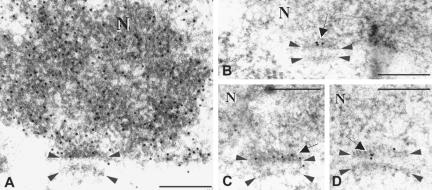
During the extended prophase to the meiosis I division, chromosomes assemble axial elements (AE) along replicated sister chromatids whose ends attach to the inner nuclear membrane (NM) via a specialized conical thickening. Here, we show at the EM level that in Sycp3(-/-) spermatocyte chromosomes lack the AE and the conical end thickening, but still they attach their telomeres to the inner NM with an electron-dense plate that contains T(2)AG(3) repeats. Immunofluorescence detected telomere proteins, SCP2, and the meiosis-specific cohesin STAG3 at the Sycp3(-/-) telomere. Bouquet stage spermatocytes were approximately threefold enriched, and the number of telomere but not centromere signals was reduced to the haploid in advanced Sycp3(-/-) spermatocytes, which indicates a special mode of homolog pairing at the mammalian telomere. Fluorescence in situ hybridization with mouse chromosome 8- and 12-specific subsatellite probes uncovered reduced levels of regional homolog pairing, whereas painting of chromosomes 13 revealed partial or complete juxtapositioning of homologs; however, condensation of Sycp3(-/-) bivalents was defective. Electron microscopic analysis of AE-deficient spermatocytes revealed that transverse filaments formed short structures reminiscent of the synaptonemal complex central region, which likely mediate stable homolog pairing. It appears that the AE is required for chromosome condensation, rapid exit from the bouquet stage, and fine-tuning of homolog pairing.
Figures



Similar articles
-
Centromere and telomere redistribution precedes homologue pairing and terminal synapsis initiation during prophase I of cattle spermatogenesis.Cytogenet Cell Genet. 2001;93(3-4):304-14. doi: 10.1159/000057002. Cytogenet Cell Genet. 2001. PMID: 11528130
-
Centromere and telomere movements during early meiotic prophase of mouse and man are associated with the onset of chromosome pairing.J Cell Biol. 1996 Sep;134(5):1109-25. doi: 10.1083/jcb.134.5.1109. J Cell Biol. 1996. PMID: 8794855 Free PMC article.
-
Transmembrane protein Sun2 is involved in tethering mammalian meiotic telomeres to the nuclear envelope.Proc Natl Acad Sci U S A. 2007 May 1;104(18):7426-31. doi: 10.1073/pnas.0609198104. Epub 2007 Apr 23. Proc Natl Acad Sci U S A. 2007. PMID: 17452644 Free PMC article.
-
Centromere Associations in Meiotic Chromosome Pairing.Annu Rev Genet. 2015;49:95-114. doi: 10.1146/annurev-genet-112414-055107. Epub 2015 Sep 29. Annu Rev Genet. 2015. PMID: 26421510 Review.
-
Knockout mice provide novel insights into meiotic chromosome and telomere dynamics.Cytogenet Genome Res. 2003;103(3-4):235-44. doi: 10.1159/000076809. Cytogenet Genome Res. 2003. PMID: 15051944 Review.
Cited by
-
Poly(ADP-ribose) polymerase-2 contributes to the fidelity of male meiosis I and spermiogenesis.Proc Natl Acad Sci U S A. 2006 Oct 3;103(40):14854-9. doi: 10.1073/pnas.0604252103. Epub 2006 Sep 25. Proc Natl Acad Sci U S A. 2006. PMID: 17001008 Free PMC article.
-
A novel mouse synaptonemal complex protein is essential for loading of central element proteins, recombination, and fertility.PLoS Genet. 2011 May;7(5):e1002088. doi: 10.1371/journal.pgen.1002088. Epub 2011 May 26. PLoS Genet. 2011. PMID: 21637789 Free PMC article.
-
Meiotic chromosome dynamics and double strand break formation in reptiles.Front Cell Dev Biol. 2022 Oct 12;10:1009776. doi: 10.3389/fcell.2022.1009776. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36313577 Free PMC article.
-
Non-homologous chromosome pairing and crossover formation in haploid rice meiosis.Chromosoma. 2011 Feb;120(1):47-60. doi: 10.1007/s00412-010-0288-3. Epub 2010 Aug 13. Chromosoma. 2011. PMID: 20706730
-
Meiosis in mice without a synaptonemal complex.PLoS One. 2011;6(12):e28255. doi: 10.1371/journal.pone.0028255. Epub 2011 Dec 2. PLoS One. 2011. PMID: 22164254 Free PMC article.
References
-
- Alsheimer, M., and Benavente, R. (1996). Change of karyoskeleton during mammalian spermatogenesis: expression pattern of nuclear lamin C2 and its regulation. Exp. Cell Res. 228, 181-188. - PubMed
-
- Barlow, A.L., and Hultén, M.A. (1997). Combined immunocytogenetic and molecular cytogenetic analysis of meiosis I oocytes from normal human females. Zygote 6, 27-38. - PubMed
-
- Bass, H.W., Bordoli, S.J., and Foss, E.M. (2003). The desynaptic (dy) and desynaptic1 (dsy1) mutations in maize (Zea mays L.) cause distinct telomere-misplacement phenotypes during meiotic prophase. J. Exp. Bot. 54, 39-46. - PubMed
-
- Benavente, R., Alsheimer, M., and von Glasenapp, E. (2004). The nuclear envelope at the attachment sites of mammalian meiotic telomeres. Chrom. Today 14 (in press).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
