

doi: 10.1093/emboj/cdg589.
Decatenation of DNA circles by FtsK-dependent Xer site-specific recombination
Affiliations
- PMID: 14633998
- PMCID: PMC291834
- DOI: 10.1093/emboj/cdg589
Item in Clipboard
Decatenation of DNA circles by FtsK-dependent Xer site-specific recombination
EMBO J.
.
Abstract
DNA replication results in interlinked (catenated) sister duplex molecules as a consequence of the intertwined helices that comprise duplex DNA. DNA topoisomerases play key roles in decatenation. We demonstrate a novel, efficient and directional decatenation process in vitro, which uses the combination of the Escherichia coli XerCD site-specific recombination system and a protein, FtsK, which facilitates simple synapsis of dif recombination sites during its translocation along DNA. We propose that the FtsK-XerCD recombination machinery, which converts chromosomal dimers to monomers, may also function in vivo in removing the final catenation links remaining upon completion of DNA replication.
Figures
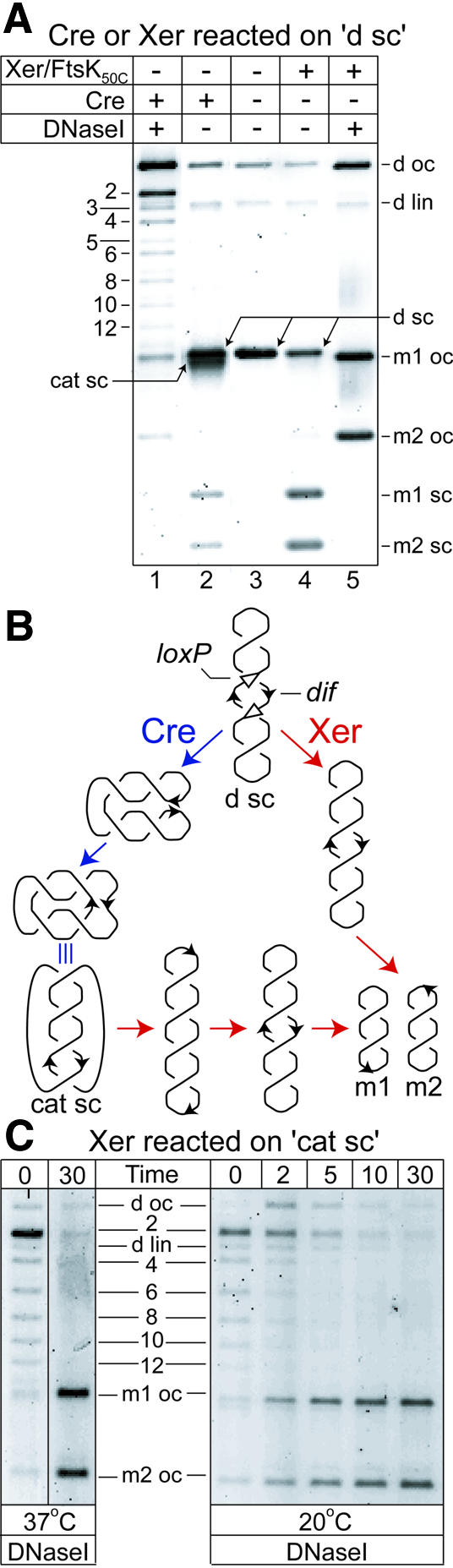
Fig. 1. FtsK50C–XerCD–dif recombination unlinks catenated circles containing antiparallel dif sites. (A) Topology of the products of Cre and FtsK50C–XerCD recombination on the 6 kb pseudo-dimeric deletion substrate, d sc (B), in which the pairs of adjacent dif and loxP sites are separated by 3.5 and 2.5 kb. The topological complexity of products is revealed after DNase I nicking (lanes 1 and 5). DNA bands are labelled: d sc, d oc and d lin, supercoiled, open circle and linear pseudo-dimeric plasmid, respectively; m1 oc and m2 oc, monomer open circles; m1 sc and m2 sc, monomer supercoils; cat sc, supercoiled catenanes; 2, 4, 6, 8, 10, 12, nicked catenanes with two, four, six, eight, 10 and 12 nodes, respectively; 3 and 5, nicked knots with three and five nodes. A small amount of HJ runs between 2-cat and 3-knot (see Figure 3A). (B) Schematic of Cre–loxP and FtsK50C–XerCD–dif recombination on the pseudo-dimeric supercoiled deletion substrate, d sc, and of FtsK50C–Xer recombination on supercoiled catenanes (cat sc). For simplicity, adjacent loxP and dif sites are shown as a single black arrow in all DNA molecules other than the initial d sc substrate. Cre–loxP and FtsK50C–XerCD–dif recombination pathways are delineated by blue and red arrows, respectively. (C) FtsK50C–Xer recombination on supercoiled catenanes containing antiparallel dif sites. Supercoiled catenanes (cat sc) were generated by Cre–loxP recombination on the supercoiled deletion substrate, d sc.
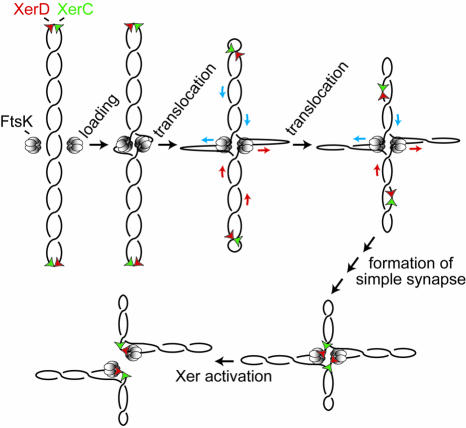
Fig. 2. One scheme by which FtsK50C (hexameric rings) might promote synapsis of directly repeated dif sites (red and green triangles) on a supercoiled DNA molecule, prior to activation of Xer recombination. Two units of FtsK50C form a double ring on DNA, by analogy with RuvB and other DNA translocases. Translocation by each ring reels in the distant dif sites and facilitates their synapsis and activation of recombination. We propose that once a XerCD-bound dif site interacts with a ring, its DNA translocation activity stalls. Models in which a single ring facilitates synapsis of dif sites are also possible, although the reaction is not as facile as that presented here. DNA translocation will generate separate positively and negatively supercoiled domains when two distant DNA segments interact simultaneously with a ring (Aussel et al., 2002). For simplicity, this differential supercoiling is not shown.
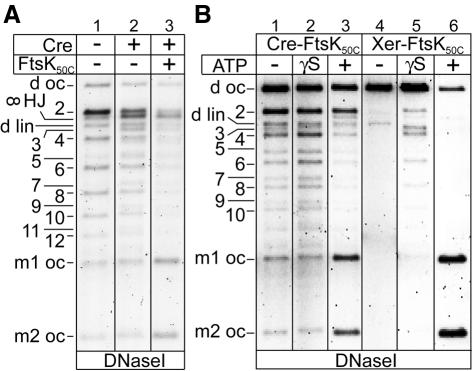
Fig. 3. FtsK50C promotes formation of free circles in recombination of plasmid dimers and catenated monomers by Cre–loxP as well as by XerCD–dif. (A) Effect of FtsK50C on Cre–loxP recombination of catenanes with antiparallel loxP sites. Samples were nicked with DNase I to reveal product topology. ∞ HJ, ‘figure 8’ nicked HJ intermediate; 2, 4, 6, 8, 10, 12, nicked catenanes with two, four, six, eight, 10 and 12 nodes, respectively; 3, 5, 7, 9 and 11, nicked knots with three, five, seven, nine and 11 nodes. (B) Influence of nucleotide on FtsK50C action during Cre–loxP and XerCD–dif recombination on the supercoiled deletion substrate, d sc (Figure 1B). γS is ATP-γS.
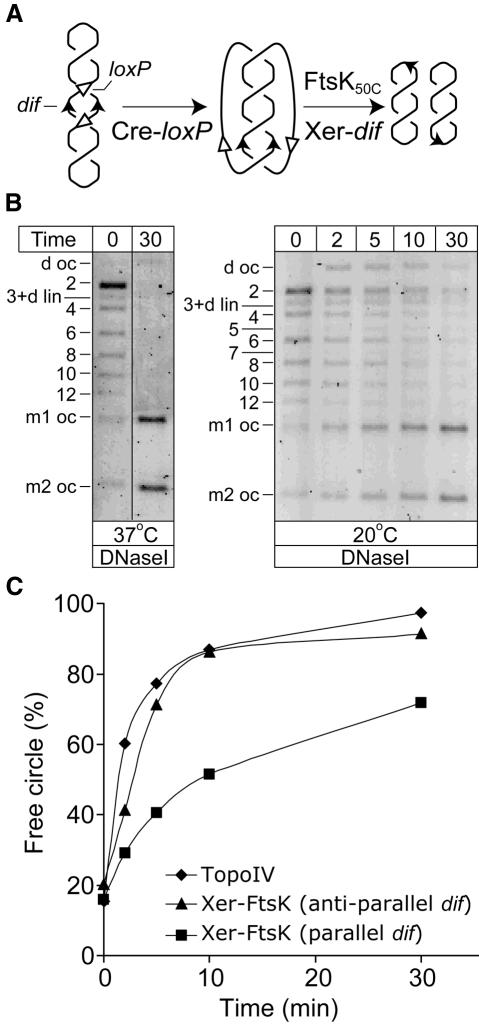
Fig. 4. Decatenation of pseudo-replicative catenanes. (A) Schematic of catenanes with parallel dif sites formed by Cre recombination at directly repeated loxP sites, and their subsequent decatenation by FtsK50C–XerCD-mediated recombination at dif. The initial plasmid was constructed in a similar way to that described in Figure 1A, except that the dif sites were in inverted repeat with respect to each other. Therefore, the catenanes generated by Cre–loxP recombination have parallel dif sites and mimic replicative catenanes. (B) FtsK50C–XerCD recombination on catenanes containing parallel dif sites. The abbreviations are as in Figure 1A. (C) Comparison of decatenation by FtsK50C–XerCD and TopoIV at 20°C. Note that only a single FtsK50C–XerCD-dif decatenating unit can be active on each molecule, whereas multiple TopoIV molecules could be active.
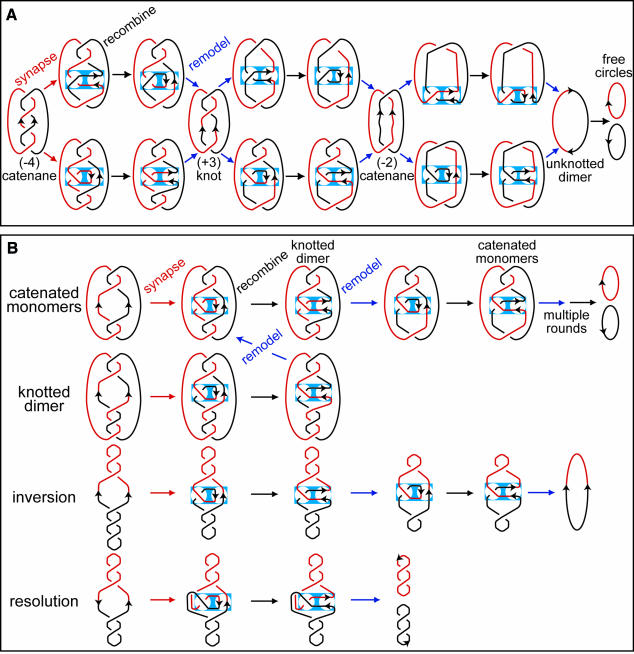
Fig. 5. Schematic of FtsK50C–XerCD-dif-mediated decatenation. (A) Comparison of alternative schemes for decatenation of pseudo-replicative catenanes by sequential rounds of Xer recombination. The consequences of the two schemes are identical, with a linking change (ΔLk) of +2 at each recombination round being accommodated through loss of one catenation–knot node and loss of one (–) supercoil. In the pathway shown at the top, a pre-existing (–) interdomainal catenation crossing is trapped in the recombination complex, and the recombination reaction generates an intradomainal (+) loop containing a single recombination site, which can cancel out a pre-existing (–) supercoil (not shown). In the pathway shown at the bottom, an intradomainal (–) supercoil loop containing a single recombination site is trapped into the recombination complex; this (–) loop can be derived from (–) plectonemic supercoiling elsewhere in the molecule (not shown). The blue rectangle represents the recombination complex formed by the XerCD heterotetramer on the synapsed dif sites. The recombination complex is portioned into two separate regions (circles). One contains the antiparallel- oriented dif sites and is changed by recombination (right-hand side), while the other is unchanged by recombination and contains the trapped (–) crossing (left-hand side). (B) Scheme showing recombination pathways in which an intradomainal supercoil is trapped into the recombination complex for a variety of recombination substrates. Decatenation, unknotting, inversion and deletion are compared. DNA contained within the recombination complex is the same in all cases. The two separate circles within the recombination complex correspond to the ‘parental (P) tangle’ (right-hand side) and ‘bound (Ob) tangle’ (left-hand side) of Crisona et al. (1999). In the configuration shown, tangle P = ∞ and recombinant tangle, R = 0; i.e the orientation of the synapsed dif sites is changed from vertical to horizontal by recombination. The (–) supercoil loop corresponding to Ob is assumed to be recruited from plectonemic supercoiling elsewhere in the molecule (only shown for the inversion substrate). Any other topological nodes are removed by FtsKC action as it facilitates synapsis. If any such nodes had been trapped, they would be incorporated into the product topology. The ∞ tangle for antiparallel recombination sites has been shown to be the most appropriate recombination site configuration for λ Int reactions, and the additional crossing bound by recombinase has been shown to be a (–) topological node for λ Int, Cre and Flp (Crisona et al., 1999; Grainge et al., 2000). The recombination sites are in exactly the same configuration before and after recombination during decatenation, unknotting, simple dimer resolution and inversion. Note that, in the plasmid deletion reaction, in addition to the (–) node trapped in the recombination complex, there is a compensatory (+) node outside the complex; this is required to maintain the correct connectivity and to ensure the products are free circles. The other way to ensure free circles during deletion is not to trap any crossings into the recombination complex. During decatenation–unknotting, each round of recombination leads to the loss of one catenation or knotting node and one (–) supercoil from within the catenane–knot, consistent with a ΔLk of +2, which has been shown experimentally for λ Int, Cre and Flp inversion.
Similar articles
-
Delayed activation of Xer recombination at dif by FtsK during septum assembly in Escherichia coli.Mol Microbiol. 2008 May;68(4):1018-28. doi: 10.1111/j.1365-2958.2008.06212.x. Epub 2008 Mar 19. Mol Microbiol. 2008. PMID: 18363794
-
Extent of the activity domain and possible roles of FtsK in the Escherichia coli chromosome terminus.Mol Microbiol. 2005 Jun;56(6):1539-48. doi: 10.1111/j.1365-2958.2005.04633.x. Mol Microbiol. 2005. PMID: 15916604
-
Unlinking chromosome catenanes in vivo by site-specific recombination.EMBO J. 2007 Oct 3;26(19):4228-38. doi: 10.1038/sj.emboj.7601849. Epub 2007 Sep 6. EMBO J. 2007. PMID: 17805344 Free PMC article.
-
Simple topology: FtsK-directed recombination at the dif site.Biochem Soc Trans. 2013 Apr;41(2):595-600. doi: 10.1042/BST20120299. Biochem Soc Trans. 2013. PMID: 23514160 Review.
-
The Escherichia coli DNA translocase FtsK.Biochem Soc Trans. 2010 Apr;38(2):395-8. doi: 10.1042/BST0380395. Biochem Soc Trans. 2010. PMID: 20298190 Review.
Cited by
-
Functional taxonomy of bacterial hyperstructures.Microbiol Mol Biol Rev. 2007 Mar;71(1):230-53. doi: 10.1128/MMBR.00035-06. Microbiol Mol Biol Rev. 2007. PMID: 17347523 Free PMC article. Review.
-
Comparative genomics of the FtsK-HerA superfamily of pumping ATPases: implications for the origins of chromosome segregation, cell division and viral capsid packaging.Nucleic Acids Res. 2004 Oct 5;32(17):5260-79. doi: 10.1093/nar/gkh828. Print 2004. Nucleic Acids Res. 2004. PMID: 15466593 Free PMC article.
-
The bacterial cell cycle, chromosome inheritance and cell growth.Nat Rev Microbiol. 2019 Aug;17(8):467-478. doi: 10.1038/s41579-019-0212-7. Nat Rev Microbiol. 2019. PMID: 31164753 Review.
-
Viral and cellular SOS-regulated motor proteins: dsDNA translocation mechanisms with divergent functions.Cell Biosci. 2014 Jun 25;4:31. doi: 10.1186/2045-3701-4-31. eCollection 2014. Cell Biosci. 2014. PMID: 24995125 Free PMC article. Review.
-
Large chromosomal rearrangements during a long-term evolution experiment with Escherichia coli.mBio. 2014 Sep 9;5(5):e01377-14. doi: 10.1128/mBio.01377-14. mBio. 2014. PMID: 25205090 Free PMC article.
References
-
- Abremski K., Hoess,R. and Sternberg,N. (1983) Studies on the properties of P1 site-specific recombination: evidence for topologically unlinked products following recombination. Cell, 32, 1301–1311. - PubMed
-
- Abremski K., Frommer,B. and Hoess,R.H. (1986) Linking-number changes in the DNA substrate during Cre-mediated loxP site-specific recombination. J. Mol. Biol., 192, 17–26. - PubMed
-
- Adams D.E., Shekhtman,E.M., Zechiedrich,E.L., Schmid,M.B. and Cozzarelli,N.R. (1992) The role of topoisomerase IV in partitioning bacterial replicons and the structure of catenated intermediates in DNA replication. Cell, 71, 277–288. - PubMed
-
- Aussel L., Barre,F.X., Aroyo,M., Stasiak,A., Stasiak,A.Z. and Sherratt,D. (2002) FtsK is a DNA motor protein that activates chromosome dimer resolution by switching the catalytic state of the XerC and XerD recombinases. Cell, 108, 195–205. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
