

Immunoglobulin somatic hypermutation: double-strand DNA breaks, AID and error-prone DNA repair
- PMID: 12959216
- PMCID: PMC4624321
- DOI: 10.1023/a:1024571714867
Immunoglobulin somatic hypermutation: double-strand DNA breaks, AID and error-prone DNA repair
Abstract
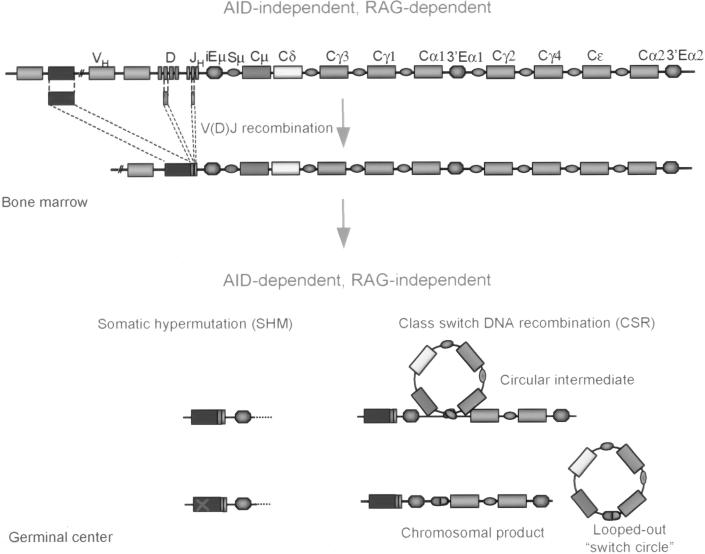
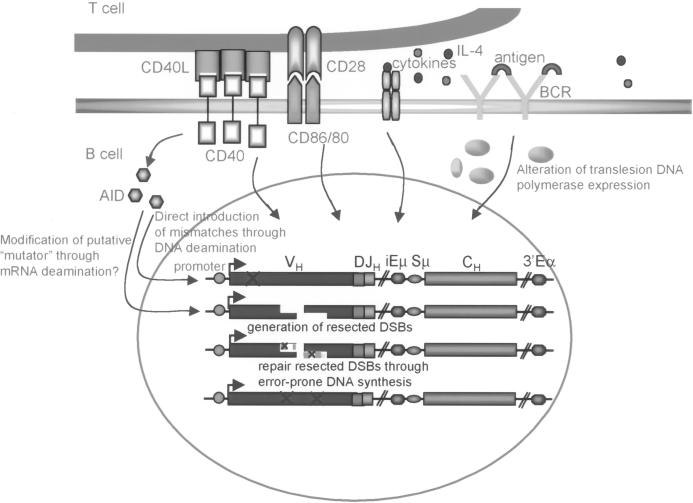
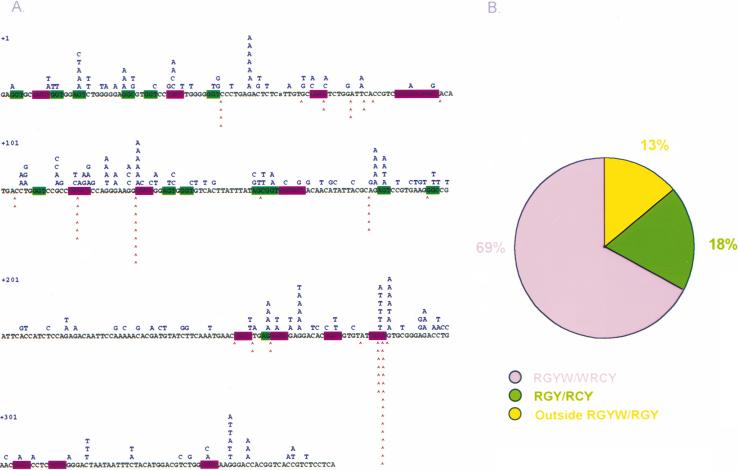
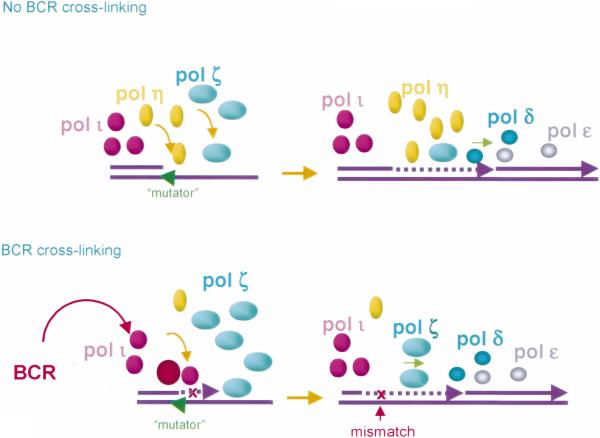
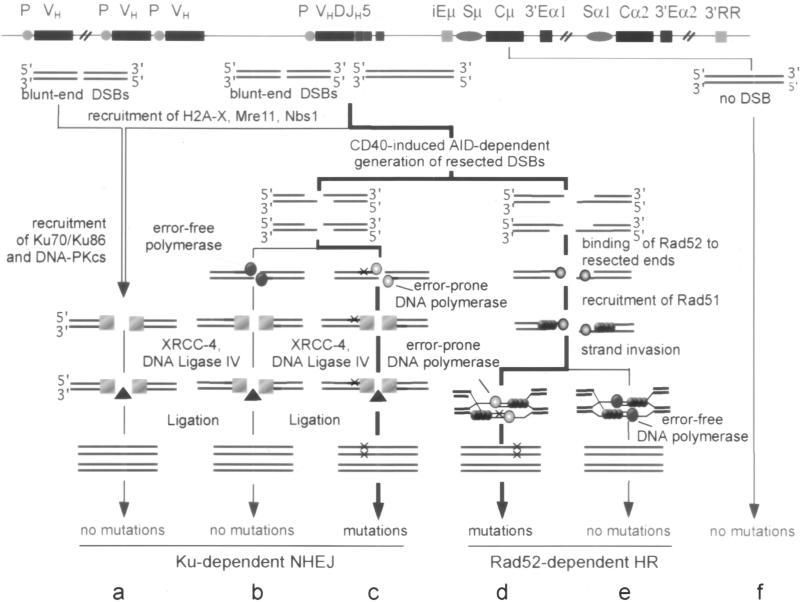
Somatic hypermutation (SHM) is critical for antibody affinity maturation and the generation of memory B cells. Somatic mutations consist mainly of single nucleotide changes with rare insertions and deletions. Such changes would be introduced during error-prone repair of lesions involving single-strand DNA breaks (SSBs) or, more likely, double-strand DNA breaks (DSBs), as DSBs occur exclusively in genes that have the potentials to undergo SHM. In the human, such genes include Ig V, BCL6, and c-MYC. In these germline genes, DSBs are blunt. In rearranged Ig V, BCL6, and translocated c-MYC genes, blunt DSBs are processed to yield resected DNA ends. This process is dependent on the expression of activation-induced cytidine deaminase (AID), which is selectively expressed upon CD40-signaling in hypermutating B cells. CD40-induced and AID-dependent free 5'- and 3'-staggered DNA ends critically channel the repair of DSBs through the homologous recombination (HR) repair pathway. During HR, the modulation of critical translesion DNA polymerases, as signaled by cross-linking of the B cell receptor (BCR) for antigen, leads to the insertions of mismatches, i.e., mutations. The nature of DSBs, the possible roles of AID in the modification of DSBs and that of the translesion DNA polymerases zeta and iota in the subsequent repair process that lead to the insertions of mutations are discussed here within the context of an integrated model of SHM.
Figures
Similar articles
-
AID- and Ung-dependent generation of staggered double-strand DNA breaks in immunoglobulin class switch DNA recombination: a post-cleavage role for AID.Mol Immunol. 2008 Nov;46(1):45-61. doi: 10.1016/j.molimm.2008.07.003. Epub 2008 Aug 28. Mol Immunol. 2008. PMID: 18760480 Free PMC article.
-
AID-dependent generation of resected double-strand DNA breaks and recruitment of Rad52/Rad51 in somatic hypermutation.Immunity. 2003 Jun;18(6):727-38. doi: 10.1016/s1074-7613(03)00151-1. Immunity. 2003. PMID: 12818155 Free PMC article.
-
DNA double strand breaks occur independent of AID in hypermutating Ig genes.Clin Dev Immunol. 2003 Jun-Dec;10(2-4):83-9. doi: 10.1080/10446670310001626571. Clin Dev Immunol. 2003. PMID: 14768938 Free PMC article.
-
DNA lesions and repair in immunoglobulin class switch recombination and somatic hypermutation.Ann N Y Acad Sci. 2005 Jun;1050:146-62. doi: 10.1196/annals.1313.119. Ann N Y Acad Sci. 2005. PMID: 16014529 Free PMC article. Review.
-
Mutating for Good: DNA Damage Responses During Somatic Hypermutation.Front Immunol. 2019 Mar 12;10:438. doi: 10.3389/fimmu.2019.00438. eCollection 2019. Front Immunol. 2019. PMID: 30915081 Free PMC article. Review.
Cited by
-
Biased dA/dT somatic hypermutation as regulated by the heavy chain intronic iEmu enhancer and 3'Ealpha enhancers in human lymphoblastoid B cells.Mol Immunol. 2006 Apr;43(11):1817-26. doi: 10.1016/j.molimm.2005.10.018. Epub 2006 Jan 10. Mol Immunol. 2006. PMID: 16412510 Free PMC article.
-
DNA repair in antibody somatic hypermutation.Trends Immunol. 2006 Jul;27(7):313-21. doi: 10.1016/j.it.2006.05.001. Epub 2006 Jun 5. Trends Immunol. 2006. PMID: 16737852 Free PMC article. Review.
-
A novel human B cell subpopulation representing the initial germinal center population to express AID.Blood. 2007 Mar 15;109(6):2545-52. doi: 10.1182/blood-2006-07-037150. Epub 2006 Nov 28. Blood. 2007. PMID: 17132718 Free PMC article.
-
Mutagenic and recombinagenic responses to defective DNA polymerase delta are facilitated by the Rev1 protein in pol3-t mutants of Saccharomyces cerevisiae.Genetics. 2008 Aug;179(4):1795-806. doi: 10.1534/genetics.108.089821. Genetics. 2008. PMID: 18711219 Free PMC article.
-
AID- and Ung-dependent generation of staggered double-strand DNA breaks in immunoglobulin class switch DNA recombination: a post-cleavage role for AID.Mol Immunol. 2008 Nov;46(1):45-61. doi: 10.1016/j.molimm.2008.07.003. Epub 2008 Aug 28. Mol Immunol. 2008. PMID: 18760480 Free PMC article.
References
-
- Pulendran B, Kannourakis G, Nouri S, Smith KG, Nossal GJ. Soluble antigen can cause enhanced apoptosis of germinal-centre B cells. Nature. 1995;375:331–334. - PubMed
-
- Cornall RJ, Goodnow CC, Cyster JG. The regulation of self-reactive B cells. Curr Opin Immunol. 1995;7:804–811. - PubMed
-
- Hertz M, Nemazee D. Receptor editing and commitment in B lymphocytes. Curr Opin Immunol. 1998;10:208–213. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials