

Gating currents associated with intramembrane charge displacement in HERG potassium channels
- PMID: 12928493
- PMCID: PMC193596
- DOI: 10.1073/pnas.1832721100
Gating currents associated with intramembrane charge displacement in HERG potassium channels
Abstract
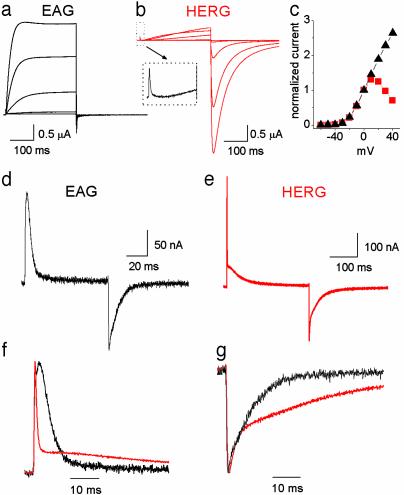
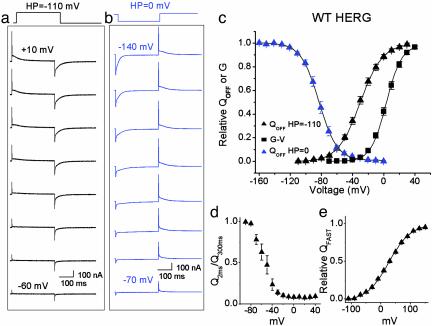
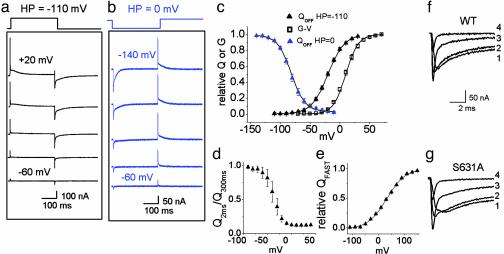
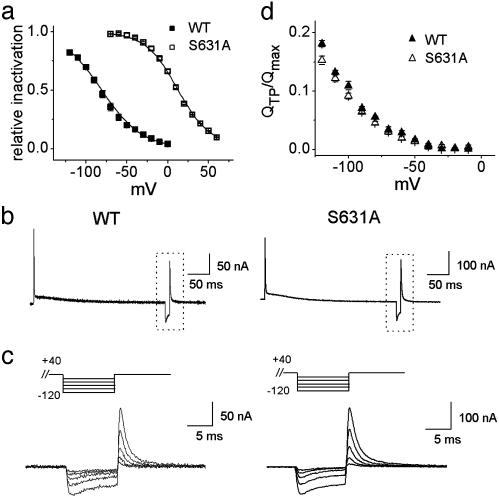
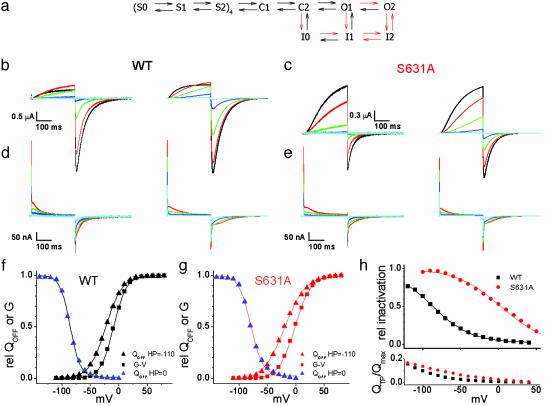
HERG (human ether-a-go-go-related gene) encodes a delayed rectifier K+ channel vital to normal repolarization of cardiac action potentials. Attenuation of repolarizing K+ current caused by mutations in HERG or channel block by common medications prolongs ventricular action potentials and increases the risk of arrhythmia and sudden death. The critical role of HERG in maintenance of normal cardiac electrical activity derives from its unusual gating properties. Opposite to other voltage-gated K+ channels, the rate of HERG channel inactivation is faster than activation and appears to be intrinsically voltage dependent. To investigate voltage sensor movement associated with slow activation and fast inactivation, we characterized HERG gating currents. When the cut-open oocyte voltage clamp technique was used, membrane depolarization elicited gating current with fast and slow components that differed 100-fold in their kinetics. Unlike previously studied voltage-gated K+ channels, the bulk of charge movement in HERG was protracted, consistent with the slow rate of ionic current activation. Despite similar kinetic features, fast inactivation was not derived from the fast gating component. Analysis of an inactivation-deficient mutant HERG channel and a Markov kinetic model suggest that HERG inactivation is coupled to activation.
Figures
Similar articles
-
Fast and slow voltage sensor movements in HERG potassium channels.J Gen Physiol. 2002 Mar;119(3):275-93. doi: 10.1085/jgp.20028534. J Gen Physiol. 2002. PMID: 11865022 Free PMC article.
-
Altered gating of HERG potassium channels by cobalt and lanthanum.Pflugers Arch. 2000 Jun;440(2):264-74. doi: 10.1007/s004240000263. Pflugers Arch. 2000. PMID: 10898527
-
Voltage sensor movement in the hERG K+ channel.Novartis Found Symp. 2005;266:46-52; discussion 52-6, 95-9. doi: 10.1002/047002142x.ch5. Novartis Found Symp. 2005. PMID: 16050261 Review.
-
Saxitoxin is a gating modifier of HERG K+ channels.J Gen Physiol. 2003 Jun;121(6):583-98. doi: 10.1085/jgp.200308812. J Gen Physiol. 2003. PMID: 12771193 Free PMC article.
-
The HERG K+ channel: progress in understanding the molecular basis of its unusual gating kinetics.Eur Biophys J. 2004 Apr;33(2):89-97. doi: 10.1007/s00249-003-0338-3. Epub 2003 Sep 10. Eur Biophys J. 2004. PMID: 13680209 Review.
Cited by
-
Computational cardiology and risk stratification for sudden cardiac death: one of the grand challenges for cardiology in the 21st century.J Physiol. 2016 Dec 1;594(23):6893-6908. doi: 10.1113/JP272015. Epub 2016 Jun 9. J Physiol. 2016. PMID: 27060987 Free PMC article.
-
Gating charges in the activation and inactivation processes of the HERG channel.J Gen Physiol. 2004 Dec;124(6):703-18. doi: 10.1085/jgp.200409119. Epub 2004 Nov 15. J Gen Physiol. 2004. PMID: 15545400 Free PMC article.
-
Molecular mapping of a site for Cd2+-induced modification of human ether-à-go-go-related gene (hERG) channel activation.J Physiol. 2005 Sep 15;567(Pt 3):737-55. doi: 10.1113/jphysiol.2005.089094. Epub 2005 Jun 23. J Physiol. 2005. PMID: 15975984 Free PMC article.
-
A gating charge interaction required for late slow inactivation of the bacterial sodium channel NavAb.J Gen Physiol. 2013 Sep;142(3):181-90. doi: 10.1085/jgp.201311012. J Gen Physiol. 2013. PMID: 23980192 Free PMC article.
-
The isolated voltage sensing domain of the Shaker potassium channel forms a voltage-gated cation channel.Elife. 2016 Oct 6;5:e18130. doi: 10.7554/eLife.18130. Elife. 2016. PMID: 27710769 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
