

PAUSED encodes the Arabidopsis exportin-t ortholog
- PMID: 12913168
- PMCID: PMC184644
- DOI: 10.1104/pp.103.023309
PAUSED encodes the Arabidopsis exportin-t ortholog
Abstract
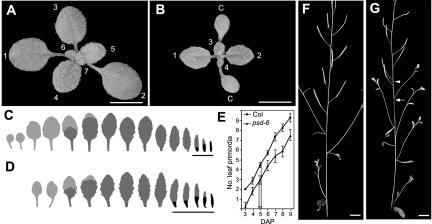
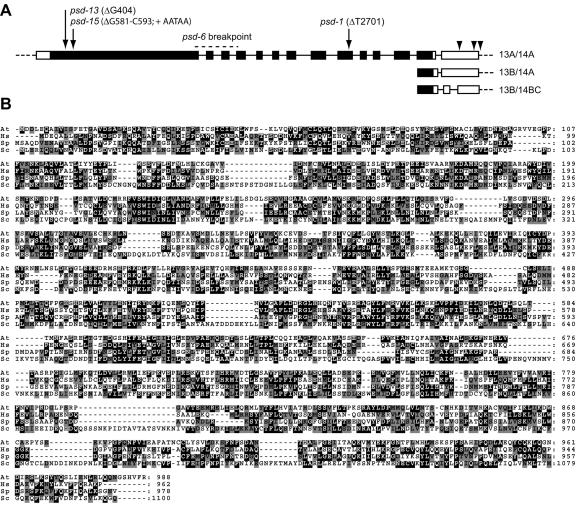
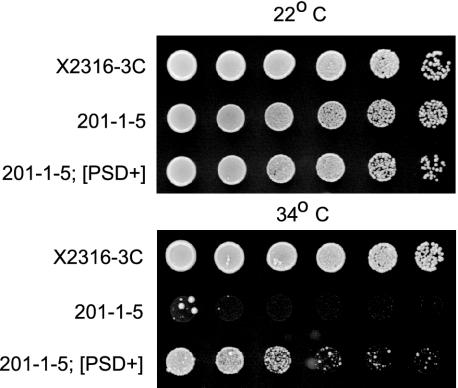
Los1p/exportin-t (XPOT) mediates the nuclear export of tRNAs in yeast and mammals. The requirements for this transport pathway are unclear, however, because los1 mutations do not affect yeast growth, and the phenotype of XPOT mutations in mammals is unknown. Here, we show that PAUSED (PSD) is the Arabidopsis ortholog of LOS1/XPOT and is capable of rescuing the tRNA export defect of los1 in Brewer's yeast (Saccharomyces cerevisiae), suggesting that its function has been conserved. Putative null alleles of PSD disrupt the initiation of the shoot apical meristem and delay leaf initiation after germination, the emergence of the radicle and lateral roots, and the transition to flowering. Plants doubly mutant for psd and hasty, the Arabidopsis ortholog of exportin 5, are viable but have a more severe phenotype than either single mutant. These results suggest that PSD plays a role in tRNA export in Arabidopsis, but that at least one-and perhaps several-additional tRNA export pathways also exist. The PSD transcript is broadly expressed during development and is alternatively spliced in the 3'-untranslated region. No temporal or spatial difference in the abundance of different splice forms was observed. We propose that the mutant phenotype of psd reflects defects in developmental events and cell/tissue types that require elevated levels of protein synthesis and are therefore acutely sensitive to a reduction in tRNA export.
Figures



Similar articles
-
PAUSED, a putative exportin-t, acts pleiotropically in Arabidopsis development but is dispensable for viability.Plant Physiol. 2003 Aug;132(4):1913-24. doi: 10.1104/pp.103.023291. Plant Physiol. 2003. PMID: 12913148 Free PMC article.
-
Expression and characterization of rice putative PAUSED gene.Acta Biochim Biophys Sin (Shanghai). 2008 Oct;40(10):893-900. Acta Biochim Biophys Sin (Shanghai). 2008. PMID: 18850055
-
HASTY, the Arabidopsis ortholog of exportin 5/MSN5, regulates phase change and morphogenesis.Development. 2003 Apr;130(8):1493-504. doi: 10.1242/dev.00362. Development. 2003. PMID: 12620976
-
ULTRAPETALA1 encodes a SAND domain putative transcriptional regulator that controls shoot and floral meristem activity in Arabidopsis.Development. 2005 Mar;132(5):897-911. doi: 10.1242/dev.01642. Epub 2005 Jan 26. Development. 2005. PMID: 15673576
-
Exportin t and Exportin 5: tRNA and miRNA biogenesis - and beyond.Biol Chem. 2012 Jul;393(7):599-604. doi: 10.1515/hsz-2012-0146. Biol Chem. 2012. PMID: 22944664 Review.
Cited by
-
Three tRNA nuclear exporters in S. cerevisiae: parallel pathways, preferences, and precision.Nucleic Acids Res. 2022 Sep 23;50(17):10140-10152. doi: 10.1093/nar/gkac754. Nucleic Acids Res. 2022. PMID: 36099418 Free PMC article.
-
Nucleo-cytoplasmic transport of proteins and RNA in plants.Plant Cell Rep. 2011 Feb;30(2):153-76. doi: 10.1007/s00299-010-0928-3. Epub 2010 Oct 20. Plant Cell Rep. 2011. PMID: 20960203 Free PMC article. Review.
-
PAUSED, a putative exportin-t, acts pleiotropically in Arabidopsis development but is dispensable for viability.Plant Physiol. 2003 Aug;132(4):1913-24. doi: 10.1104/pp.103.023291. Plant Physiol. 2003. PMID: 12913148 Free PMC article.
-
Rice Importin β1 gene affects pollen tube elongation.Mol Cells. 2011 Jun;31(6):523-30. doi: 10.1007/s10059-011-2321-7. Epub 2011 Apr 14. Mol Cells. 2011. PMID: 21499832 Free PMC article.
-
tRNA Processing and Subcellular Trafficking Proteins Multitask in Pathways for Other RNAs.Front Genet. 2019 Feb 20;10:96. doi: 10.3389/fgene.2019.00096. eCollection 2019. Front Genet. 2019. PMID: 30842788 Free PMC article. Review.
References
-
- Arts GJ, Fornerod M, Mattaj IW (1998a) Identification of a nuclear export receptor for tRNA. Curr Biol 8: 305-314 - PubMed
-
- Barton MK, Poethig RS (1993) Formation of the shoot apical meristem in Arabidopsis thaliana and analysis of development in the wild type and in the shoot meristemless mutant. Development 119: 823-831
-
- Blazquez MA, Soowal LN, Lee I, Weigel D (1997) LEAFY expression and flower initiation in Arabidopsis. Development 124: 3835-3844 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
