

State-dependent processing of sensory stimuli by thalamic reticular neurons
- PMID: 12832551
- PMCID: PMC6741167
- DOI: 10.1523/JNEUROSCI.23-12-05264.2003
State-dependent processing of sensory stimuli by thalamic reticular neurons
Abstract
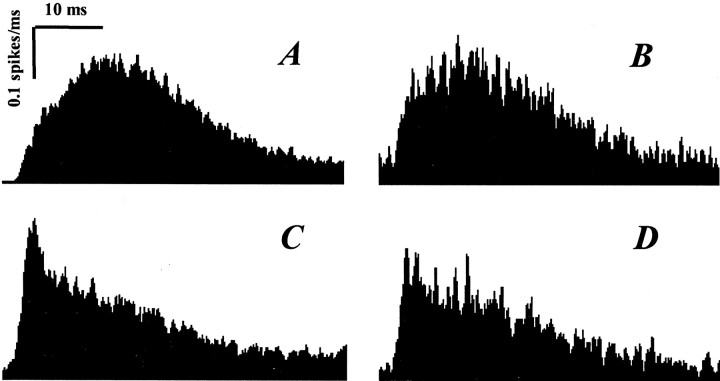
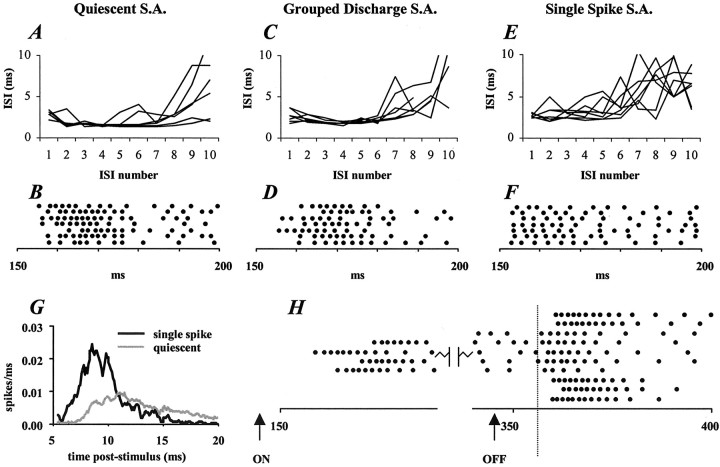
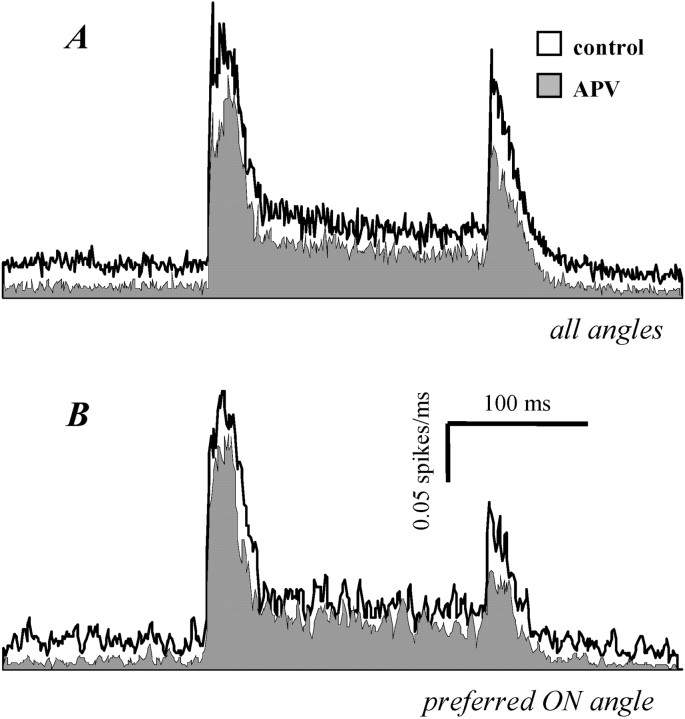
Inhibitory neurons of the thalamic reticular (RT) nucleus fire in two activity modes, burst and tonic, depending on an animal's behavioral state. In tonic mode, depolarized RT cells fire single action potentials continuously, whereas burst firing consists of grouped discharges separated by periods of quiescence. To determine how these firing modes affect sensory-evoked RT responses, single-unit responses to controlled whisker deflections were analyzed according to the burst versus tonic mode of spontaneous activity (SA) preceding the response. After burst mode activity (i.e., either quiescence or spontaneous bursts), responses exhibited a slow approximately 15 msec rise to peak firing rates followed by a approximately 35 msec decay. Interspike intervals within the response exhibited accelerando-decelerando patterns similar to those of spontaneous bursts. After tonic mode activity (i.e., single spikes), responses had a nearly instantaneous approximately 1.5 msec rise-to-peak followed by a approximately 40 msec decay, with large spike counts (5.2 spikes per stimulus) similar to those evoked in burst mode (6.2 spikes per stimulus). Interspike intervals were longer in tonic mode and exhibited a decelerando pattern. Initial evoked spikes, however, had shorter latencies and greater synchrony, contributing to the rapid onset of tonic population response. Shifts from quiescent (presumed burst mode) to tonic SA could be induced by either previous whisker deflections or iontophoretic application of NMDA; both manipulations effected appropriate shifts from burst to tonic response spike patterns. In awake animals, burst and tonic firing in RT, as in thalamocortical relay nuclei, may reflect sensory processing strategies appropriate for different behavioral and attentional states.
Figures




Similar articles
-
Inhibition suppresses transmission of tonic vibrissa-evoked activity in the rat ventrobasal thalamus.J Neurosci. 2000 Oct 1;20(19):RC100. doi: 10.1523/JNEUROSCI.20-19-j0001.2000. J Neurosci. 2000. PMID: 11000200 Free PMC article.
-
Mechanisms for signal transformation in lemniscal auditory thalamus.J Neurophysiol. 1996 Dec;76(6):3597-608. doi: 10.1152/jn.1996.76.6.3597. J Neurophysiol. 1996. PMID: 8985860
-
Effects of membrane voltage on receptive field properties of lateral geniculate neurons in the cat: contributions of the low-threshold Ca2+ conductance.J Neurophysiol. 1992 Dec;68(6):2185-98. doi: 10.1152/jn.1992.68.6.2185. J Neurophysiol. 1992. PMID: 1337104
-
Spike timing and synaptic dynamics at the awake thalamocortical synapse.Prog Brain Res. 2005;149:91-105. doi: 10.1016/S0079-6123(05)49008-1. Prog Brain Res. 2005. PMID: 16226579 Review.
-
Tonic and burst firing: dual modes of thalamocortical relay.Trends Neurosci. 2001 Feb;24(2):122-6. doi: 10.1016/s0166-2236(00)01714-8. Trends Neurosci. 2001. PMID: 11164943 Review.
Cited by
-
Circuits formultisensory integration and attentional modulation through the prefrontal cortex and the thalamic reticular nucleus in primates.Rev Neurosci. 2007;18(6):417-38. doi: 10.1515/revneuro.2007.18.6.417. Rev Neurosci. 2007. PMID: 18330211 Free PMC article. Review.
-
Thalamic reticular nucleus induces fast and local modulation of arousal state.Elife. 2015 Oct 13;4:e08760. doi: 10.7554/eLife.08760. Elife. 2015. PMID: 26460547 Free PMC article.
-
Early Thalamic Injury After Resuscitation From Severe Asphyxial Cardiac Arrest in Developing Rats.Front Cell Dev Biol. 2021 Dec 7;9:737319. doi: 10.3389/fcell.2021.737319. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34950655 Free PMC article.
-
Dynamic corticothalamic modulation of the somatosensory thalamocortical circuit during wakefulness.Nat Commun. 2024 Apr 25;15(1):3529. doi: 10.1038/s41467-024-47863-8. Nat Commun. 2024. PMID: 38664415 Free PMC article.
-
Schizophrenia: from neurophysiological abnormalities to clinical symptoms.Front Psychol. 2015 Apr 20;6:478. doi: 10.3389/fpsyg.2015.00478. eCollection 2015. Front Psychol. 2015. PMID: 25941510 Free PMC article. No abstract available.
References
-
- Cox CL, Huguenard JR, Prince DA ( 1996) Heterogeneous axonal arborizations of rat thalamic reticular neurons in the ventrobasal nucleus. J Comp Neurol 366: 416–430. - PubMed
-
- Dworkin BR, Dworkin S ( 1995) Learning of physiological responses: II. Classical conditioning of the baroreflex. Behav Neurosci 109: 1119–1136. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources