

Pyramidal cell communication within local networks in layer 2/3 of rat neocortex
- PMID: 12813147
- PMCID: PMC2343144
- DOI: 10.1113/jphysiol.2003.044784
Pyramidal cell communication within local networks in layer 2/3 of rat neocortex
Abstract
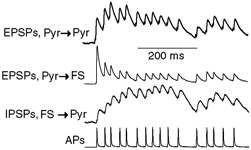
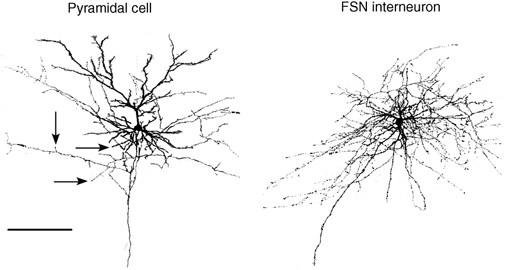
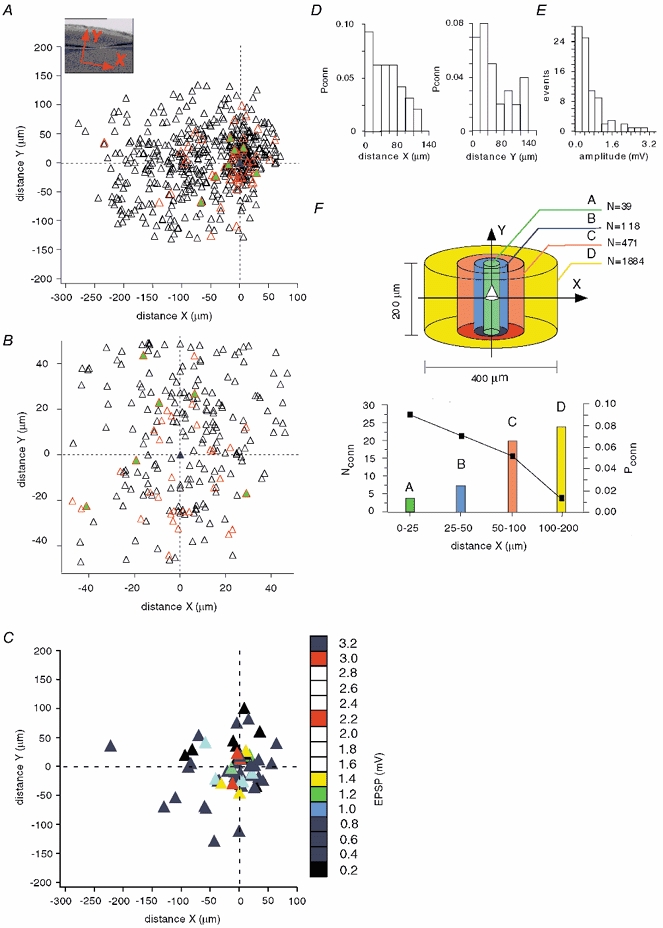
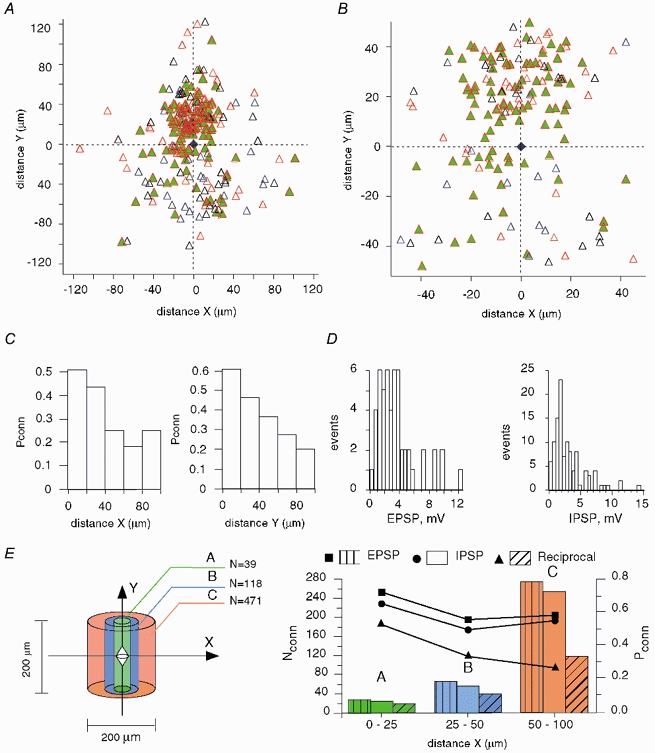
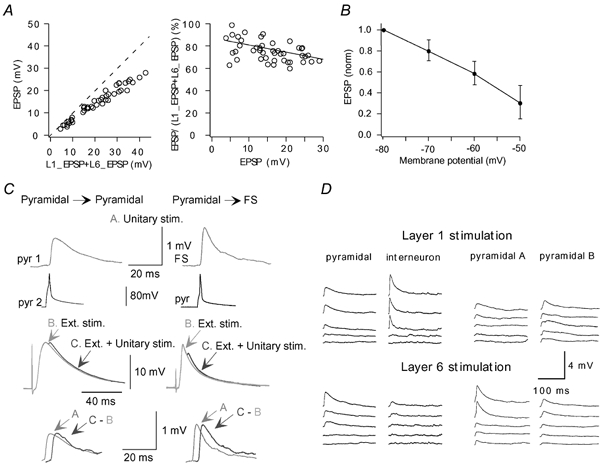
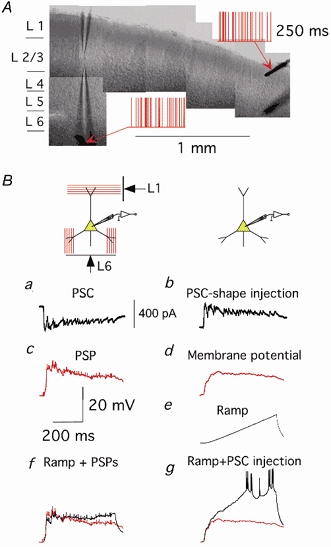
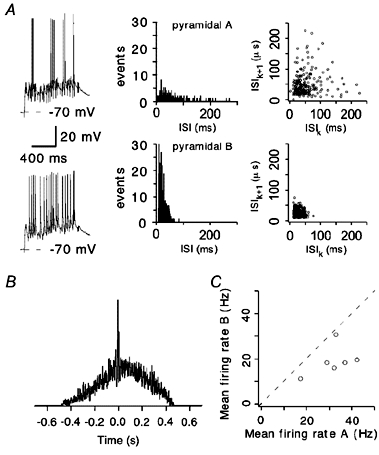
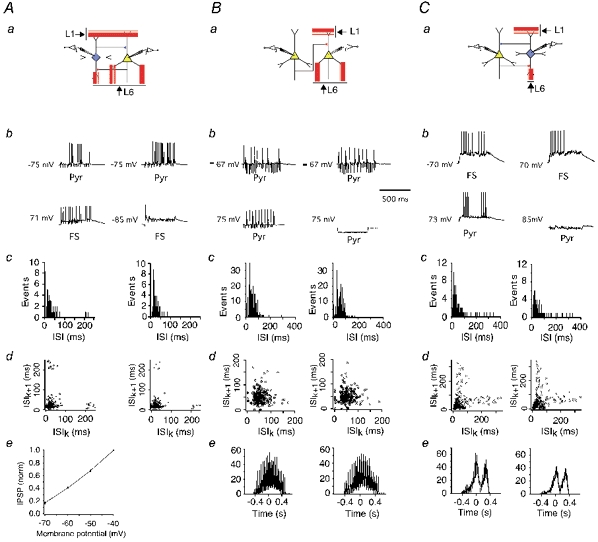
The extent to which neocortical pyramidal cells function as a local network is determined by the strength and probability of their connections. By mapping connections between pyramidal cells we show here that in a local network of about 600 pyramidal cells located within a cylindrical volume of 200 microm x 200 microm of neocortical layer 2/3, an individual pyramidal cell receives synaptic inputs from about 30 other pyramidal neurons, with the majority of EPSP amplitudes in the 0.2-1.0 mV range. The probability of connection decreased from 0.09 to 0.01 with intercell distance (over the range 25-200 microm). Within the same volume, local interneuron (fast-spiking non-accommodating interneuron, FS)-pyramidal cell connections were about 10 times more numerous, with the majority of connections being reciprocal. The probability of excitatory and inhibitory connections between pyramidal cells and FS interneurons decreased only slightly with distance, being in the range 0.5-0.75. Pyramidal cells in the local network received strong synaptic input during stimulation of afferent fibres in layers 1 and 6. Minimal-like stimulation of layer 1 or layer 6 inputs simultaneously induced postsynaptic potentials in connected pyramidal cells as well as in pyramidal-FS cell pairs. These inputs readily induced firing of pyramidal cells, although synaptically connected cells displayed different firing patterns. Unitary EPSPs in pyramidal-pyramidal cell pairs did not detectably alter cell firing. FS interneurons fire simultaneously with pyramidal cells. In pyramidal-FS cell pairs, both unitary EPSPs and IPSPs efficiently modulated cell firing patterns. We suggest that computation in the local network may proceed not only by direct pyramidal-pyramidal cell communication but also via local interneurons. With such a high degree of connectivity with surrounding pyramidal cells, local interneurons are ideally poised to both coordinate and expand the local pyramidal cell network via pyramidal-interneuron-pyramidal communication.
Figures
Similar articles
-
Distinct Ca2+ channels mediate transmitter release at excitatory synapses displaying different dynamic properties in rat neocortex.Cereb Cortex. 2006 Mar;16(3):386-93. doi: 10.1093/cercor/bhi117. Epub 2005 May 25. Cereb Cortex. 2006. PMID: 15917483
-
Efficacy and connectivity of intracolumnar pairs of layer 2/3 pyramidal cells in the barrel cortex of juvenile rats.J Physiol. 2006 Sep 1;575(Pt 2):583-602. doi: 10.1113/jphysiol.2006.105106. Epub 2006 Jun 22. J Physiol. 2006. PMID: 16793907 Free PMC article.
-
Distinct local circuits between neocortical pyramidal cells and fast-spiking interneurons in young adult rats.J Neurophysiol. 2003 Feb;89(2):943-53. doi: 10.1152/jn.00750.2002. J Neurophysiol. 2003. PMID: 12574470
-
Anatomical and functional differentiation of glutamatergic synaptic innervation in the neocortex.J Physiol Paris. 1999 Sep-Oct;93(4):305-17. doi: 10.1016/s0928-4257(00)80059-5. J Physiol Paris. 1999. PMID: 10574120 Review.
-
Interlaminar connections in the neocortex.Cereb Cortex. 2003 Jan;13(1):5-14. doi: 10.1093/cercor/13.1.5. Cereb Cortex. 2003. PMID: 12466210 Review.
Cited by
-
Frequency selective neuronal modulation triggers spreading depolarizations in the rat endothelin-1 model of stroke.J Cereb Blood Flow Metab. 2021 Oct;41(10):2756-2768. doi: 10.1177/0271678X211013656. Epub 2021 May 9. J Cereb Blood Flow Metab. 2021. PMID: 33969731 Free PMC article.
-
Feed-Forward Propagation of Temporal and Rate Information between Cortical Populations during Coherent Activation in Engineered In Vitro Networks.Front Neural Circuits. 2016 Apr 22;10:32. doi: 10.3389/fncir.2016.00032. eCollection 2016. Front Neural Circuits. 2016. PMID: 27147977 Free PMC article.
-
A neuromorphic implementation of multiple spike-timing synaptic plasticity rules for large-scale neural networks.Front Neurosci. 2015 May 20;9:180. doi: 10.3389/fnins.2015.00180. eCollection 2015. Front Neurosci. 2015. PMID: 26041985 Free PMC article.
-
Coding principles of the canonical cortical microcircuit in the avian brain.Proc Natl Acad Sci U S A. 2015 Mar 17;112(11):3517-22. doi: 10.1073/pnas.1408545112. Epub 2015 Feb 17. Proc Natl Acad Sci U S A. 2015. PMID: 25691736 Free PMC article.
-
Effective Suppression of Pathological Synchronization in Cortical Networks by Highly Heterogeneous Distribution of Inhibitory Connections.Front Comput Neurosci. 2016 Oct 18;10:109. doi: 10.3389/fncom.2016.00109. eCollection 2016. Front Comput Neurosci. 2016. PMID: 27803659 Free PMC article.
References
-
- Borg-Graham LJ, Monier C, Fregnac Y. Visual input evokes transient and strong shunting inhibition in visual cortical neurons. Nature. 1998;393:369–373. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources