

Activation of the tumour suppressor kinase LKB1 by the STE20-like pseudokinase STRAD
- PMID: 12805220
- PMCID: PMC162144
- DOI: 10.1093/emboj/cdg292
Activation of the tumour suppressor kinase LKB1 by the STE20-like pseudokinase STRAD
Abstract
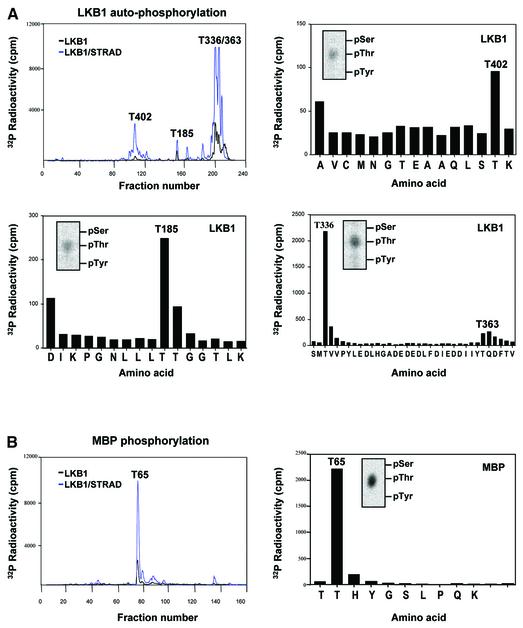
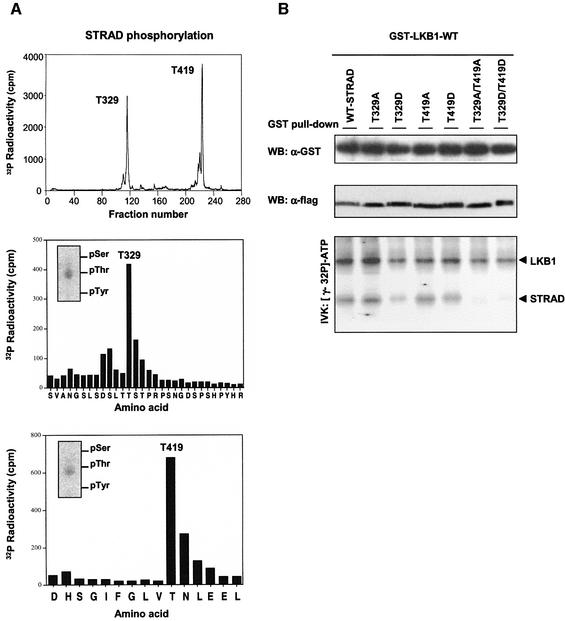
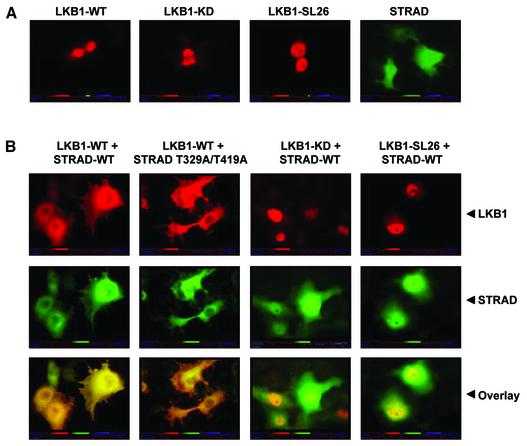
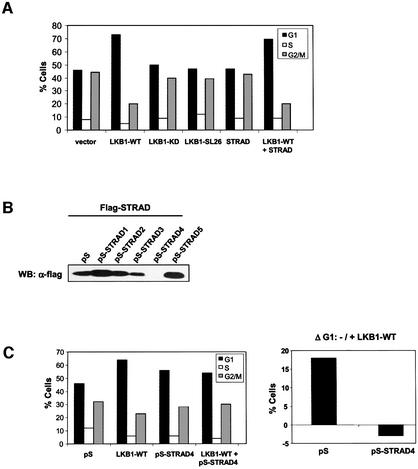
The LKB1 gene encodes a serine/threonine kinase mutated in Peutz-Jeghers cancer syndrome. Despite several proposed models for LKB1 function in development and in tumour suppression, the detailed molecular action of LKB1 remains undefined. Here, we report the identification and characterization of an LKB1-specific adaptor protein and substrate, STRAD (STe20 Related ADaptor). STRAD consists of a STE20- like kinase domain, but lacks several residues that are indispensable for intrinsic catalytic activity. Endogenous LKB1 and STRAD form a complex in which STRAD activates LKB1, resulting in phosphorylation of both partners. STRAD determines the subcellular localization of wild-type, but not mutant LKB1, translocating it from nucleus to cytoplasm. One LKB1 mutation previously identified in a Peutz-Jeghers family that does not compromise its kinase activity is shown here to interfere with LKB1 binding to STRAD, and hence with STRAD-dependent regulation. Removal of endogenous STRAD by siRNA abrogates the LKB1-induced G(1) arrest. Our results imply that STRAD plays a key role in regulating the tumour suppressor activities of LKB1.
Figures
Similar articles
-
Analysis of the LKB1-STRAD-MO25 complex.J Cell Sci. 2004 Dec 15;117(Pt 26):6365-75. doi: 10.1242/jcs.01571. Epub 2004 Nov 23. J Cell Sci. 2004. PMID: 15561763
-
Complexes between the LKB1 tumor suppressor, STRAD alpha/beta and MO25 alpha/beta are upstream kinases in the AMP-activated protein kinase cascade.J Biol. 2003;2(4):28. doi: 10.1186/1475-4924-2-28. Epub 2003 Sep 24. J Biol. 2003. PMID: 14511394 Free PMC article.
-
The ubiquitin-associated domain of AMPK-related kinases regulates conformation and LKB1-mediated phosphorylation and activation.Biochem J. 2006 Mar 15;394(Pt 3):545-55. doi: 10.1042/BJ20051844. Biochem J. 2006. PMID: 16396636 Free PMC article.
-
LKB1-dependent signaling pathways.Annu Rev Biochem. 2006;75:137-63. doi: 10.1146/annurev.biochem.75.103004.142702. Annu Rev Biochem. 2006. PMID: 16756488 Review.
-
LKB1 biology: assessing the therapeutic relevancy of LKB1 inhibitors.Cell Commun Signal. 2024 Jun 6;22(1):310. doi: 10.1186/s12964-024-01689-5. Cell Commun Signal. 2024. PMID: 38844908 Free PMC article. Review.
Cited by
-
LKB1 controls human bronchial epithelial morphogenesis through p114RhoGEF-dependent RhoA activation.Mol Cell Biol. 2013 Jul;33(14):2671-82. doi: 10.1128/MCB.00154-13. Epub 2013 May 6. Mol Cell Biol. 2013. PMID: 23648482 Free PMC article.
-
SNRK: a metabolic regulator with multifaceted role in development and disease.Vessel Plus. 2020;4:26. Epub 2020 Aug 21. Vessel Plus. 2020. PMID: 32968716 Free PMC article.
-
Dynamic Lkb1-TORC1 signaling as a possible mechanism for regulating the endoderm-intestine transition.Dev Dyn. 2010 Nov;239(11):3000-12. doi: 10.1002/dvdy.22437. Dev Dyn. 2010. PMID: 20925120 Free PMC article.
-
Regulation of gene expression in the intestinal epithelium.Prog Mol Biol Transl Sci. 2010;96:207-29. doi: 10.1016/B978-0-12-381280-3.00009-9. Prog Mol Biol Transl Sci. 2010. PMID: 21075346 Free PMC article. Review.
-
Loss of LKB1 disrupts breast epithelial cell polarity and promotes breast cancer metastasis and invasion.J Exp Clin Cancer Res. 2014 Sep 2;33(1):70. doi: 10.1186/s13046-014-0070-0. J Exp Clin Cancer Res. 2014. PMID: 25178656 Free PMC article.
References
-
- Bardeesy N., Sinha,M., Hezel,A.F., Signoretti,S., Hathaway,N.A., Sharpless,N.E., Loda,M., Carrasco,D.R. and DePinho,R.A. (2002) Loss of the Lkb1 tumour suppressor provokes intestinal polyposis but resistance to transformation. Nature, 419, 162–167. - PubMed
-
- Brummelkamp T.R., Bernards,R. and Agami,R. (2002) A system for stable expression of short interfering RNAs in mammalian cells. Science, 296, 550–552. - PubMed
-
- Candotti F. et al. (1997) Structural and functional basis for JAK3-deficient severe combined immunodeficiency. Blood, 90, 3996–4003. - PubMed
-
- Castrop J., van Wichen,D., Koomans-Bitter,M., Van de,W.M., de Weger,R., van Dongen,J. and Clevers,H. (1995) The human TCF-1 gene encodes a nuclear DNA-binding protein uniquely expressed in normal and neoplastic T-lineage lymphocytes. Blood, 86, 3050–3059. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
