

Immune responses to RNA-virus infections of the CNS
- PMID: 12776209
- PMCID: PMC7097089
- DOI: 10.1038/nri1105
Immune responses to RNA-virus infections of the CNS
Abstract
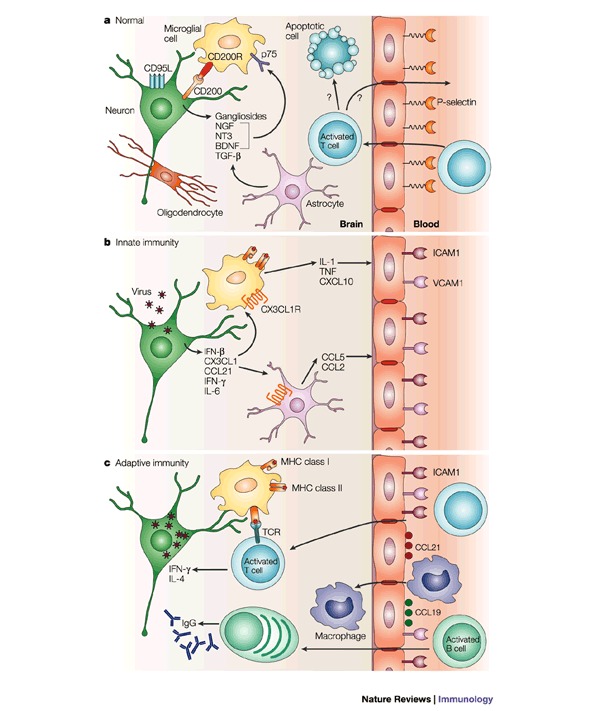
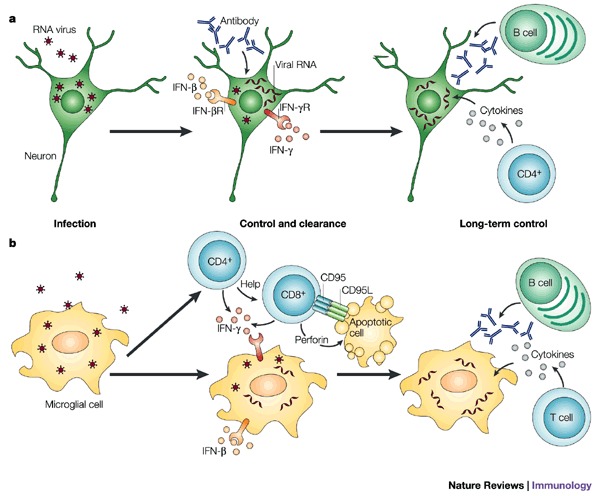
A successful outcome for the host of virus infection of the central nervous system (CNS) requires the elimination of the virus without damage to essential non-renewable cells, such as neurons. As a result, inflammatory responses must be tightly controlled, and many unique mechanisms seem to contribute to this control. In addition to being important causes of human disease, RNA viruses that infect the CNS provide useful models in which to study immune responses in the CNS. Recent work has shown the importance of innate immune responses in the CNS in controlling virus infection. And advances have been made in assessing the relative roles of cytotoxic T cells, antibodies and cytokines in the clearance of viruses from neurons, glial cells and meningeal cells.
Figures
Similar articles
-
Keeping it in check: chronic viral infection and antiviral immunity in the brain.Nat Rev Neurosci. 2016 Dec;17(12):766-776. doi: 10.1038/nrn.2016.140. Epub 2016 Nov 4. Nat Rev Neurosci. 2016. PMID: 27811921 Free PMC article. Review.
-
Alpha-Synuclein Expression Restricts RNA Viral Infections in the Brain.J Virol. 2015 Dec 30;90(6):2767-82. doi: 10.1128/JVI.02949-15. J Virol. 2015. PMID: 26719256 Free PMC article.
-
Germ Line IgM Is Sufficient, but Not Required, for Antibody-Mediated Alphavirus Clearance from the Central Nervous System.J Virol. 2018 Mar 14;92(7):e02081-17. doi: 10.1128/JVI.02081-17. Print 2018 Apr 1. J Virol. 2018. PMID: 29321331 Free PMC article.
-
MHV infection of the CNS: mechanisms of immune-mediated control.Viral Immunol. 2001;14(1):1-18. doi: 10.1089/08828240151061329. Viral Immunol. 2001. PMID: 11270593 Review.
-
Regulatory T cells selectively preserve immune privilege of self-antigens during viral central nervous system infection.J Immunol. 2012 Apr 15;188(8):3678-85. doi: 10.4049/jimmunol.1102422. Epub 2012 Mar 9. J Immunol. 2012. PMID: 22407917
Cited by
-
Virus-specific and bystander CD8 T cells recruited during virus-induced encephalomyelitis.J Virol. 2005 Apr;79(8):4700-8. doi: 10.1128/JVI.79.8.4700-4708.2005. J Virol. 2005. PMID: 15795256 Free PMC article.
-
Immunological thresholds in neurological gene therapy: highly efficient elimination of transduced cells might be related to the specific formation of immunological synapses between T cells and virus-infected brain cells.Neuron Glia Biol. 2006 Nov;2(4):309-22. doi: 10.1017/S1740925X07000579. Neuron Glia Biol. 2006. PMID: 18084640 Free PMC article.
-
Attenuation of Activated eIF2α Signaling by ISRIB Treatment After Spinal Cord Injury Improves Locomotor Function.J Mol Neurosci. 2022 Mar;72(3):585-597. doi: 10.1007/s12031-021-01920-9. Epub 2021 Oct 13. J Mol Neurosci. 2022. PMID: 34647267 Free PMC article.
-
IL-15 independent maintenance of virus-specific CD8(+) T cells in the CNS during chronic infection.J Neuroimmunol. 2009 Feb 15;207(1-2):32-8. doi: 10.1016/j.jneuroim.2008.11.005. Epub 2008 Dec 23. J Neuroimmunol. 2009. PMID: 19106006 Free PMC article.
-
Neuroimmunology of central nervous system viral infections: the cells, molecules and mechanisms involved.Curr Opin Pharmacol. 2008 Aug;8(4):472-9. doi: 10.1016/j.coph.2008.05.002. Epub 2008 Jun 16. Curr Opin Pharmacol. 2008. PMID: 18562249 Free PMC article. Review.
References
-
- Stevenson PG, Freeman S, Bangham CR, Hawke S. Virus dissemination through the brain parenchyma without immunologic control. J. Immunol. 1997;159:1876–1884. - PubMed
-
- Hoek RM, et al. Down-regulation of the macrophage lineage through interaction with OX2 (CD200) Science. 2000;290:1768–1771. - PubMed
-
- Johnson MD, Gold LI, Moses HL. Evidence for transforming growth factor-β expression in human leptomeningeal cells and transforming growth factor-β-like activity in human cerebrospinal fluid. Lab. Invest. 1992;67:360–368. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
