

Hyperglycosylated mutants of human immunodeficiency virus (HIV) type 1 monomeric gp120 as novel antigens for HIV vaccine design
- PMID: 12719582
- PMCID: PMC154011
- DOI: 10.1128/jvi.77.10.5889-5901.2003
Hyperglycosylated mutants of human immunodeficiency virus (HIV) type 1 monomeric gp120 as novel antigens for HIV vaccine design
Abstract
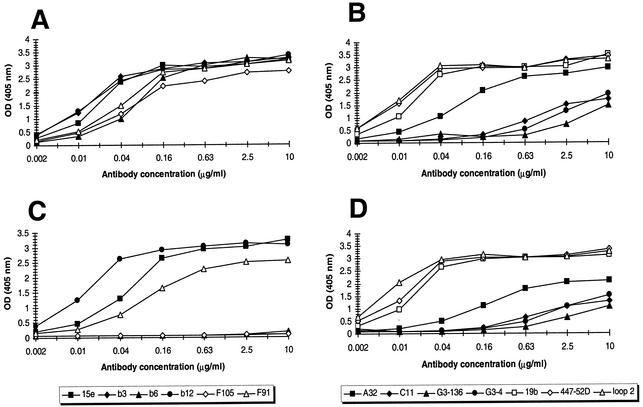
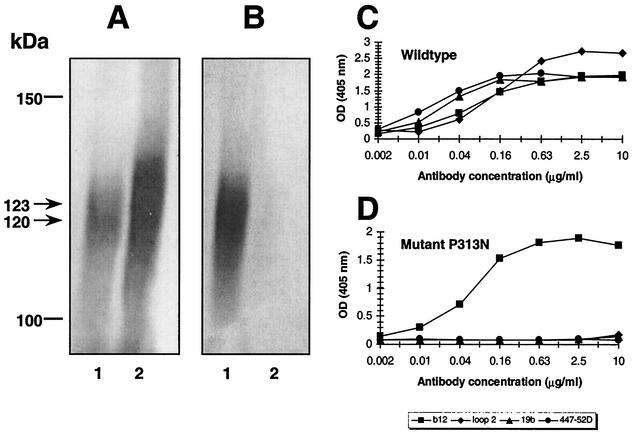
The ability to induce broadly neutralizing antibodies should be a key component of any forthcoming vaccine against human immunodeficiency virus type 1. One potential vaccine candidate, monomeric gp120, has generally failed to elicit such antibodies. We postulated that gp120 might be a better immunogen if it could be engineered to preferentially bind known broadly neutralizing antibodies. In a first study, we found that four alanine substitutions on the perimeter of the so-called Phe-43 cavity of gp120 could reduce binding of weakly neutralizing CD4-binding site antibodies (R. Pantophlet, E. O. Saphire, P. Poignard, P. W. H. I. Parren, I. A. Wilson, and D. R. Burton, J. Virol. 77:642-658, 2003), while slightly enhancing binding of the potent, broadly neutralizing antibody b12. In the present study, we sought to reduce or abolish the binding of a wider range of nonneutralizing antibodies, by incorporating extra N-glycosylation motifs at select positions into the hypervariable loops and the gp120 core. A hyperglycosylated mutant containing seven extra glycosylation sequons (consensus sequences) and the four alanine substitutions described above did not bind an extensive panel of nonneutralizing and weakly neutralizing antibodies, including a polyclonal immunoglobulin preparation (HIVIG) of low neutralizing potency. Binding of b12, at lowered affinity, and of four antibodies to the C1 and C5 regions was maintained. Removal of N- and C-terminal residues in the C1 and C5 regions, respectively, reduced or abolished binding of the four antibodies, but this also adversely affected b12 binding. The hyperglycosylated mutant and its analogues described here are novel antigens that may provide a new approach to eliciting antibodies with b12-like neutralizing properties.
Figures
Similar articles
-
Improved design of an antigen with enhanced specificity for the broadly HIV-neutralizing antibody b12.Protein Eng Des Sel. 2004 Oct;17(10):749-58. doi: 10.1093/protein/gzh085. Epub 2004 Nov 12. Protein Eng Des Sel. 2004. PMID: 15542540
-
Hyperglycosylated stable core immunogens designed to present the CD4 binding site are preferentially recognized by broadly neutralizing antibodies.J Virol. 2014 Dec;88(24):14002-16. doi: 10.1128/JVI.02614-14. Epub 2014 Sep 24. J Virol. 2014. PMID: 25253346 Free PMC article.
-
Fine mapping of the interaction of neutralizing and nonneutralizing monoclonal antibodies with the CD4 binding site of human immunodeficiency virus type 1 gp120.J Virol. 2003 Jan;77(1):642-58. doi: 10.1128/jvi.77.1.642-658.2003. J Virol. 2003. PMID: 12477867 Free PMC article.
-
GP120: target for neutralizing HIV-1 antibodies.Annu Rev Immunol. 2006;24:739-69. doi: 10.1146/annurev.immunol.24.021605.090557. Annu Rev Immunol. 2006. PMID: 16551265 Review.
-
The implications of antigenic diversity for vaccine development.Immunol Lett. 1999 Mar;66(1-3):159-64. doi: 10.1016/s0165-2478(98)00176-x. Immunol Lett. 1999. PMID: 10203049 Review.
Cited by
-
Identifying epitopes of HIV-1 that induce protective antibodies.Nat Rev Immunol. 2004 Mar;4(3):199-210. doi: 10.1038/nri1307. Nat Rev Immunol. 2004. PMID: 15039757 Free PMC article. Review.
-
Antibody responses elicited in macaques immunized with human immunodeficiency virus type 1 (HIV-1) SF162-derived gp140 envelope immunogens: comparison with those elicited during homologous simian/human immunodeficiency virus SHIVSF162P4 and heterologous HIV-1 infection.J Virol. 2006 Sep;80(17):8745-62. doi: 10.1128/JVI.00956-06. J Virol. 2006. PMID: 16912322 Free PMC article.
-
25 years of HIV research on virology, virus restriction, immunopathogenesis, genes and vaccines.Clin Exp Immunol. 2008 Oct;154(1):6-14. doi: 10.1111/j.1365-2249.2008.03750.x. Epub 2008 Aug 29. Clin Exp Immunol. 2008. PMID: 18761662 Free PMC article. Review.
-
A platform incorporating trimeric antigens into self-assembling nanoparticles reveals SARS-CoV-2-spike nanoparticles to elicit substantially higher neutralizing responses than spike alone.Sci Rep. 2020 Oct 23;10(1):18149. doi: 10.1038/s41598-020-74949-2. Sci Rep. 2020. PMID: 33097791 Free PMC article.
-
Antibody Responses Elicited by Immunization with BG505 Trimer Immune Complexes.J Virol. 2019 Sep 30;93(20):e01188-19. doi: 10.1128/JVI.01188-19. Print 2019 Oct 15. J Virol. 2019. PMID: 31375582 Free PMC article.
References
-
- Arthur, L. O., J. W. Bess, Jr., E. N. Chertova, J. L. Rossio, M. T. Esser, R. E. Benveniste, L. E. Henderson, and J. D. Lifson. 1998. Chemical inactivation of retroviral infectivity by targeting nucleocapsid protein zinc fingers: a candidate SIV vaccine. AIDS Res. Hum. Retrovir. 14(Suppl. 3):S311-S319. - PubMed
-
- Back, N. K. T., L. Smit, J.-J. de Jong, W. Keulen, M. Schutten, J. Goudsmit, and M. Tersmette. 1994. An N-glycan within the human immunodeficiency virus type 1 gp120 V3 loop affects virus neutralization. Virology 199:431-438. - PubMed
-
- Barbas, C. F., III, E. Bjorling, F. Chiodi, N. Dunlop, D. Cababa, T. M. Jones, S. L. Zebedee, M. A. Persson, P. L. Nara, E. Norrby, and D. R. Burton. 1992. Recombinant human Fab fragments neutralize human type 1 immunodeficiency virus in vitro. Proc. Natl. Acad. Sci. USA 89:9339-9343. - PMC - PubMed
-
- Barbas, C. F., III, T. A. Collet, W. Amberg, P. Roben, J. M. Binley, D. Hoekstra, D. Cababa, T. M. Jones, R. A. Williamson, G. R. Pilkington, N. L. Haigwood, E. Cabezas, A. C. Satterthwait, I. Sanz, and D. R. Burton. 1993. Molecular profile of an antibody response to HIV-1 as probed by combinatorial libraries. J. Mol. Biol. 230:812-823. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
