

Intracellular assembly and secretion of recombinant subviral particles from tick-borne encephalitis virus
- PMID: 12634393
- PMCID: PMC150630
- DOI: 10.1128/jvi.77.7.4370-4382.2003
Intracellular assembly and secretion of recombinant subviral particles from tick-borne encephalitis virus
Abstract
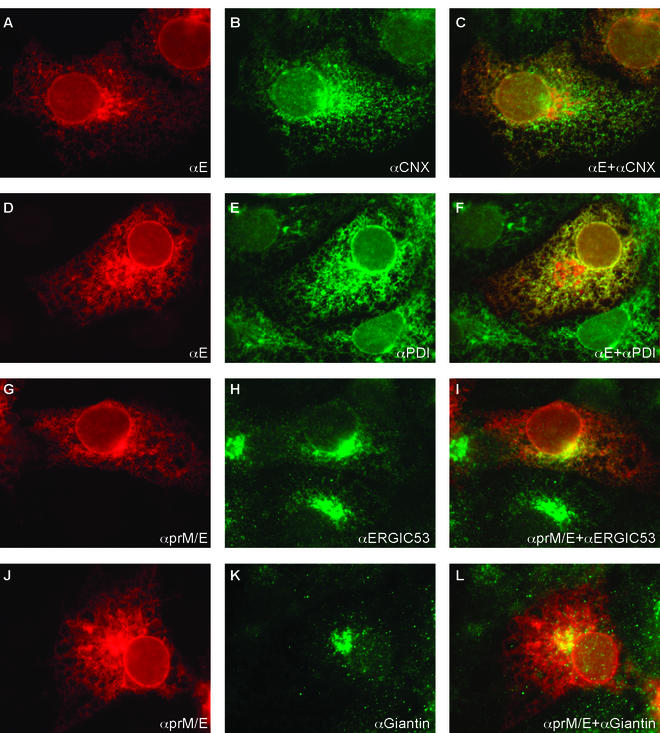
It is believed that flavivirus assembly occurs by intracellular budding of the nucleocapsid into the lumen of the endoplasmic reticulum (ER). Recombinant expression of tick-borne encephalitis (TBE) virus envelope proteins prM and E in mammalian cells leads to their incorporation into enveloped recombinant subviral particles (RSPs), which have been used as a model system for studying assembly and entry processes and are also promising vaccine candidates. In this study, we analyzed the formation and secretion of TBE virus RSPs and of a membrane anchor-free E homodimer in mammalian cells. Immunofluorescence microscopy showed that E was accumulated in the lumen of the ER. RSPs were observed by electron microscopy in the rough and smooth ER and in downstream compartments of the secretory pathway. About 75% of the particles appeared to be of the size expected for RSPs (about 30 nm in diameter), but a number of larger particles and tubular structures were also observed in these compartments. Secretion of membrane anchor-free E dimers was detected 30 min after synthesis of prM and E, and secretion of RSPs was detected 1 h after synthesis of prM and E. We also found that the presence of the single N-linked oligosaccharide side chain on the E protein and its trimming by glucosidases was necessary for secretion of RSPs and truncated E dimers. Our results suggest that incorporation of prM and E into RSPs occurs at the ER membrane without other viral elements being required, followed by rapid transport along the compartments of the secretory pathway and secretion. Moreover, the carbohydrate side chain of E is involved in at least one assembly or transport step.
Figures




Similar articles
-
Two distinct size classes of immature and mature subviral particles from tick-borne encephalitis virus.J Virol. 2003 Nov;77(21):11357-66. doi: 10.1128/jvi.77.21.11357-11366.2003. J Virol. 2003. PMID: 14557621 Free PMC article.
-
Molecular organization of a recombinant subviral particle from tick-borne encephalitis virus.Mol Cell. 2001 Mar;7(3):593-602. doi: 10.1016/s1097-2765(01)00206-4. Mol Cell. 2001. PMID: 11463384
-
Recombinant subviral particles from tick-borne encephalitis virus are fusogenic and provide a model system for studying flavivirus envelope glycoprotein functions.J Virol. 1996 Jul;70(7):4549-57. doi: 10.1128/JVI.70.7.4549-4557.1996. J Virol. 1996. PMID: 8676481 Free PMC article.
-
Molecular mechanisms of flavivirus membrane fusion.Amino Acids. 2011 Nov;41(5):1159-63. doi: 10.1007/s00726-009-0370-4. Epub 2009 Nov 1. Amino Acids. 2011. PMID: 19882217 Review.
-
Immunogenicity against Far Eastern and Siberian subtypes of tick-borne encephalitis (TBE) virus elicited by the currently available vaccines based on the European subtype: systematic review and meta-analysis.Hum Vaccin Immunother. 2014;10(10):2819-33. doi: 10.4161/hv.29984. Hum Vaccin Immunother. 2014. PMID: 25483679 Free PMC article. Review.
Cited by
-
Immunodominant West Nile Virus T Cell Epitopes Are Fewer in Number and Fashionably Late.J Immunol. 2016 May 15;196(10):4263-73. doi: 10.4049/jimmunol.1501821. Epub 2016 Apr 20. J Immunol. 2016. PMID: 27183642 Free PMC article.
-
Comprehensive N-glycosylation mapping of envelope glycoprotein from tick-borne encephalitis virus grown in human and tick cells.Sci Rep. 2020 Aug 6;10(1):13204. doi: 10.1038/s41598-020-70082-2. Sci Rep. 2020. PMID: 32764711 Free PMC article.
-
Mutations in the West Nile prM protein affect VLP and virion secretion in vitro.Virology. 2012 Nov 10;433(1):35-44. doi: 10.1016/j.virol.2012.07.011. Epub 2012 Jul 31. Virology. 2012. PMID: 22858174 Free PMC article.
-
Uncoupling cis-Acting RNA elements from coding sequences revealed a requirement of the N-terminal region of dengue virus capsid protein in virus particle formation.J Virol. 2012 Jan;86(2):1046-58. doi: 10.1128/JVI.05431-11. Epub 2011 Nov 9. J Virol. 2012. PMID: 22072762 Free PMC article.
-
A toggle switch controls the low pH-triggered rearrangement and maturation of the dengue virus envelope proteins.Nat Commun. 2014 May 20;5:3877. doi: 10.1038/ncomms4877. Nat Commun. 2014. PMID: 24846574 Free PMC article.
References
-
- Adams, S. C., A. K. Broom, L. M. Sammels, A. C. Hartnett, M. J. Howard, R. J. Coelen, J. S. Mackenzie, and R. A. Hall. 1995. Glycosylation and antigenic variation among Kunjin virus isolates. Virology 206:49-56. - PubMed
-
- Allison, S. L., C. W. Mandl, C. Kunz, and F. X. Heinz. 1994. Expression of cloned envelope protein genes from the flavivirus tick-borne encephalitis virus in mammalian cells and random mutagenesis by PCR. Virus Genes 8:187-198. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
